Diplopoda in Mexican Amber: A Supplementary Checklist
Diplopoda en ámbar mexicano: Listado taxonómico adicional
Álvarez-Rodríguez, Michelle1 ; Riquelme, Francisco1,2,*; Hernández-Patricio, Miguel3; Cupul-Magaña, Fabio4
; Riquelme, Francisco1,2,*; Hernández-Patricio, Miguel3; Cupul-Magaña, Fabio4
1 Centro de Investigaciones Biológicas, Universidad Autónoma del Estado de Morelos, C.P. 62209, Cuernavaca, Morelos, Mexico.
2 Maestría en Manejo de Recursos Naturales, Universidad Autónoma del Estado de Morelos, C.P. 62209, Cuernavaca, Morelos, Mexico.
3Unafiliate
4 Centro Universitario de la Costa, Universidad de Guadalajara, C.P. 48280, Puerto Vallarta, Jalisco, Mexico.
* francisco.riquelme@uaem.mx
Abstract
The fossil record of millipedes (Myriapoda: Diplopoda) in Mexico is primarily found in the amber-bearing beds of the upper strata of the Simojovel Formation in Chiapas, dating to the uppermost late Oligocene and the lowermost Early Miocene boundary, ca. 24 Ma. We present a taxonomic checklist of fossil millipedes identified in Mexico, complementing the existing global catalog. This revised compilation encompasses 85 amber inclusions, organized by the following orders: Polyxenida (five records), Platydesmida (four records), Siphonophorida (17 records), Polydesmida (49 records), Stemmiulida (one record), Julida (one record), and others unassigned due to preservation constraints. New records of Chelodesmidae, Pyrgodesmidae, Sphaeriodesmidae, and Trichopolydesmidae are included within the Polydesmida. Descriptions and comments for each record are provided. This inventory highlights the biodiversity of Diplopoda in southern North America and supports efforts to conserve fossil material in Mexico.
Keywords: Chiapas Amber-Lagerstätte, Fossil diversity, Simojovel Formation, Simojovelite.
Resumen
El registro fósil de milpiés (Myriapoda: Diplopoda) en México se encuentra principalmente en los estratos de ámbar de la parte superior de la Formación Simojovel en Chiapas, que data del límite superior del Oligoceno tardío y el inferior del Mioceno Temprano, ca. 24 Ma. Presentamos una lista taxonómica revisada de milpiés fósiles en México para complementar el catálogo global existente. Esta compilación revisada incluye 85 inclusiones de ámbar, organizadas en los siguientes órdenes: Polyxenida (cinco registros), Platydesmida (cuatro registros), Siphonophorida (17 registros), Polydesmida (49 registros), Stemmiulida (un registro), Julida (un registro) y otros no asignados debido a restricciones de preservación. Se incluyen nuevos registros de Chelodesmidae, Pyrgodesmidae, Sphaeriodesmidae, y Trichopolydesmidae dentro de Polydesmida. Se proporcionan descripciones y comentarios para cada registro. Este inventario destaca la biodiversidad de Diplopoda en el sur de Norteamérica y apoya los esfuerzos de conservación del material fósil en México.
Palabras clave: Ambar-Lagerstätte de Chiapas, diversidad fósil, Formación Simojovel, simojovelita.
1. Introductión
The class Diplopoda de Blainville in Gervais, 1844 encompasses 299 fossil records spanning from the Middle Silurian to the Upper Pleistocene. These records are categorized into three subclasses, which include two infraclasses, six superorders, 25 orders, 15 superfamilies, 55 families, 93 genera, and 161 fossil species. It is distributed across three geological eras: the Paleozoic comprises 97 species, the Mesozoic 24 species, and the Cenozoic 42 species (Álvarez-Rodríguez et al., 2024). Furthermore, the fossil specimens exhibit a variety of preservation forms, consisting of 123 impressions, 99 compressions, and 26 ichnofossils, predominantly derived from Paleozoic and Mesozoic strata. At the same time, 165 amber inclusions have been sourced from Late Mesozoic and Cenozoic deposits. Millipedes from the Cenozoic era are mostly preserved as amber inclusions in exceptional conservation sites, including the Baltic region from the late Eocene, Mexico from the late Oligocene-Early Miocene sites, India from the Early Miocene deposits, and the Dominican Republic from the Early Miocene sites (Álvarez-Rodríguez et al., 2024).
The taxonomic composition of millipedes during the Mesozoic shows an increase at the levels of orders and families. However, this trend tends to decline throughout the Cenozoic (Álvarez-Rodríguez et al., 2024). Therefore, it is expected that the Mexican fossil record from the Middle Cenozoic will exhibit less variation at the order and family levels, but an increase in diversity at lower ranks, such as genera and species (Riquelme and Hernández-Patricio, 2018).
In Mexico, the first quantification of fossil millipedes was published by Riquelme and Hernández-Patricio (2018). Subsequently, a comprehensive global compendium of fossil Diplopoda was compiled, incorporating new records from Mexico (Álvarez-Rodríguez et al., 2024). The present work aims to complement Mexico's list by providing a comprehensive inventory of millipedes preserved in Oligo-Miocene amber and supplementary descriptions and comments for each record.
2. Geological setting
The primary amber deposits in the Los Altos de Chiapas region of southwestern Mexico are found in Simojovel, Totolapa, and Estrella de Belén. These three key sites exhibit coeval sedimentary deposits and are part of the Amber-Lagerstätte chronologically situated between the uppermost late Oligocene and the lowermost Early Miocene, ca. 24–20 Ma (Riquelme et al., 2025). Amber from Simojovel originates from the uppermost part of the late Oligocene Simojovel Formation. Totolapa amber comes from the Early Miocene Totolapa Sandstone, while Estrella de Belén amber is derived from the uppermost strata of the late Oligocene Mompuyil Formation.
The amber-bearing strata predominantly consist of limestones, siltstones, shales, fine-grained fossiliferous sandstones, lignite, iron oxides, and pyrite nodules (Riquelme et al., 2025). Evidence from the sedimentary environment and associated paleobiota indicates these deposits originated in transitional settings, spanning shallow platforms and coastal environments, consistent with a tropical mangrove forest (Allison, 1967; Frost and Langenheim, 1974; Graham, 1999; Langenheim, 2003; Perrilliat et al., 2010 ; Riquelme et al., 2025). This amber, known as Simojovelite due to its mineral properties (Riquelme et al., 2014a), originates from an extinct legume species of the genus Hymenaea Linné (sensu Langenheim, 1966).
3. Material and methods
The fossil specimens examined in this study are housed at the Colección de Paleontología, Universidad Autónoma del Estado de Morelos, Cuernavaca, Morelos, Mexico. Additional specimens are housed at the Museo del Ámbar de Chiapas (MACH) in San Cristóbal de las Casas, Chiapas, Mexico; the Museo del Ámbar Lilia Mijangos (MALM) in San Cristóbal de las Casas, Chiapas, Mexico, and the Colección de Artrópodos Fósiles, Instituto Politécnico Nacional, Mexico City, Mexico. Fossil material was acquired through fieldwork initiated in 2017, primarily in collaboration with local Tzotzil miners and artisans, who provided the raw amber and specimens. Amber was cut and polished using various abrasives to enhance the visibility of the fossil inclusions within it. Photomicrographs of isolated specimens were taken using a Carl Zeiss microscope that utilizes multiple image stacking (Z ≥ 25). Taxonomic treatment follows the expanded traditional Linnaean classification ranks. Nomenclature and terminology follow Hoffman et al. (1996), Hoffman (1999), Sierwald and Bond (2007), Golovatch (2013), Koch (2015), Enghoff et al. (2015), Edgecombe (2015), and Sierwald and Spelda (2019). The checklist database is accessible on the website http://www.riquelmelab.org.mx, which is updated regularly.
Locality Abbreviations:
Hu/ER—Huitiupán, El Rosario mine.
Pa/EB—Palenque, Estrella de Belén mine.
Si/CLG—Simojovel, Campo La Granja mine.
Si/EC—Simojovel, El Chapayal mine.
Si/Pa—Simojovel, Pabuchil mine.
Si/EPo—Simojovel, El Porvenir mine.
Si/GV—Simojovel, Guadalupe Victoria mine.
Si/LP—Simojovel, Los Pocitos mine.
Si/MC—Simojovel, Monte Cristo mine.
Si/SA—Simojovel, San Antonio mine.
To/RS—Totolapa, Río Salado mine.
Repository Abbreviations:
CAF—Colección de Artrópodos Fósiles, Escuela Nacional de Ciencias Biológicas, Instituto Politécnico Nacional, Mexico City, Mexico.
CPAL-UAEM—Colección de Paleontología, Universidad Autónoma del Estado de Morelos, Morelos, Mexico.
MACH—Museo del Ámbar de Chiapas, San Cristóbal de las Casas, Chiapas, Mexico.
MALM—Museo del Ámbar Lilia Mijangos, San Cristóbal de las Casas, Chiapas, Mexico.
4. Results and discussion
4.1. Systematic paleontology
Class Diplopoda de Blainville in Gervais, 1844
Subclass Penicillata Latreille, 1831
Order Polyxenida Verhoeff, 1934
Polyxenida indet. (Figure 1a)
Referred material. three specimens, CPAL.112, CPAL.126 (Figure 1a), CPAL.143.
Locality: Si/GV (CPAL.112), Si/LP (CPAL.126), and Si/MC (CPAL.143).
Repository. CPAL-UAEM (3).
Description. Three small–bodied specimens, with uncalcified cuticle, covered by serrated setae arranged in trichomes. CPAL.112 has a trunk with 10 rings, CPAL.126 has 10 rings, and CPAL.143 has 11 rings. Following Enghoff et al. (2015), head with trochobothria, antennae with eight antennomers plus apical disc, collum reduced; tergites, pleurites, and sternites free, not fused; telson with elongated trichomes.
Comments. Polyxenida in Mexico currently includes two families, two genera, and four living species (Bueno-Villegas et al., 2004). The fossil record is known primarily from amber inclusions, including the lowermost Late Cretaceous Myanmar amber (Wesener and Moritz, 2018) and Oligo-Miocene Mexican amber (Álvarez-Rodríguez et al., 2024).
Superfamily Polyxenoidea Lucas, 1840
Family Polyxenidae Lucas, 1840
Polyxenidae indet.
Referred material. two specimens, CPAL.111, and CPAL.209.
Locality. To/RS (CPAL.111), and Si/MC (CPAL.209).
Repository. CPAL-UAEM (2).
Description. CPAL.111 and CPAL.209 have a conspicuous telson with two dense bundles of trichomes. According to Enghoff et al. (2015), adult polyxenids typically have ten tergites and 13 pairs of legs, whereas fewer are observed in CPAL.111, which has eight rings and CPAL.209; both are preliminarily considered to be juvenile.
Comments. The family Polyxenidae comprises 22 genera and 96 living species (Enghoff, 2015), with a global distribution. It is the largest of the four New World families within the subclass Penicillata and is commonly found in the Neotropics (Hoffman et al., 1996). Ten fossil species of Polyxenidae have been described in the lowermost Late Cretaceous Myanmar amber (Wesener and Moritz, 2018). Polyxenids have also been previously reported in Mexican amber (Álvarez-Rodríguez et al., 2024).
Subclass Chilognatha Latreille, 1802/1803
Infraclass Helminthomorpha Pocock, 1887
Subterclass Colobognatha Brandt, 1834
Order Platydesmida Cook, 1895
Platydesmida indet. (Figure 1b)
Referred material. four specimens, CPAL.123 ♀ (Figure 1b), CPAL.157 ♀, CPAL.161 ♀, and CPAL.207.
Locality. Si/GV (CPAL.123), Si/MC (CPAL.157, CPAL.161), and Si/SA (CPAL.207).
Repository. CPAL-UAEM (4).
Description. Four specimens with head capsule small, pyriform, with lobes projecting laterally, eyes absent, antennae slender, antennomeres thinner than wide. CPAL.123 has a trunk with 37 rings, CPAL.157 has 36 rings, CPAL.161 has 50 rings, and CPAL.207 has 20 rings. According to Enghoff (2015), all specimens have ozopores in the fifth segment, on both sides of the paranota; tergites and pleurites strongly fused, without visible suture, free sternites; tergites with well–developed dorsal suture, metazone with horizontal paranota and narrow prozonite; legs with 7 podomeros.
Comments. Platydesmida is found in North and Central America, extending as far south as Panama, as well as in Southeast Asia, which includes Japan, Korea, mainland China, and Taiwan. It is also present in Mediterranean Europe and the Caucasus (Shelley and Golovatch, 2011; Enghoff, 2015). The diversity of Platydesmida currently includes 14 genera and 63 living species (Shear, 2011). According to Hoffman et al. (1996), Platydesmida is considered endemic to Mesoamerica. As presented here, the fossil record in Mexico indicates its distribution throughout the southernmost part of North America since the end of the Oligocene to the Early Miocene, ca. 24–20 Ma. However, the fossil record of Platydesmida dates back to the lowermost Late Cretaceous amber from Myanmar, ca. 99 Ma (Wesener and Moritz, 2018).
Order Siphonophorida Newport, 1844
Siphonophorida indet. (Figure 1c-d)
Referred material. 18 specimens, CPAL.118.1, CPAL.133 (split in two parts), CPAL.142: ♀, CPAL.162, CPAL.163, CPAL.164, CPAL.165, CPAL.166, CPAL.175, CPAL.180, CPAL.203: ♀, CPAL.204, CPAL.212, CPAL.213, CPAL.218 (Figure 1c-d), CPAL.118.1, and CPAL.182.2.
Locality. Si/Pa (CPAL.180), Si/LP (CPAL.118.1, CPAL.118.2, CPAL.133, CPAL.142, CPAL.162, CPAL.163, CPAL.164, CPAL.165, CPAL.166, and CPAL.175), Si/MC (CPAL.203, CPAL.212, and CPAL.219), Si/SA (CPAL.213), PA/EB (CPAL.204).
Repository. CPAL-UAEM (18).
Description. All are long millipedes, with a slender body and densely setose cuticle. Head small, elongated, beak–shaped; antennae robust with six antennomers; tergites without dorsal suture; legs with six podomeres, tarsus always with an accessory claw. Following Enghoff et al. (2015), CPAL.118.1 has a trunk with 28 rings, CPAL.118.2 with 30 rings, CPAL.133 with 30 rings anterior part, CPAL.142 with 88 rings, CPAL.162 with 35 rings, CPAL.163 with 36 rings, CPAL.164 with 41 rings, CPAL.165 with 36 rings, CPAL.166 with 40 rings, CPAL.175 with 42 rings, CPAL.180 with 40 rings, CPAL.182 with 30 rings, CPAL.203 with 81 rings, CPAL.212 with 47 rings, CPAL.213 with 45 rings, and CPAL.218 with 32 rings.
Comments. Living representatives of the order Siphonophorida are known to be the longest millipedes. They are currently found in tropical and subtropical regions of the Americas (as far north as the United States), South Africa, Asia (as far north as Pakistan), eastern Australia, and New Zealand (Shelley and Golovatch, 2011; Enghoff, 2015). Current diversity encompasses two families, 14 genera, and 115 extant species (Enghoff, 2015). The fossil record of Siphonophorida includes two families, two genera, and three species (Álvarez-Rodríguez et al., 2024).
Subterclass Eugnatha Attems, 1898
Superorder Merochaeta Cook, 1895
Order Polydesmida Pocock, 1887
Polydesmida indet.
Referred material. 21 specimens, CPAL.125.
1, CPAL.125.2, CPAL.135, CPAL.137, CPAL.139:♀, CPAL.140, CPAL.144: ♀, CPAL.146, CPAL.149 (split in two parts), CPAL.153, CPAL.156, CPAL.172, CPAL.177, CPAL.178, CPAL.181: ♂, CPAL.183: ♀, CPAL.199, CPAL.215, CPAL.219, MACH.22, new record CAF-5: ♀.
Locality. Si/CLG (CPAL.140), Si/Pa (CPAL.178), Si/GV (CPAL.137, and CPAL.139); Si/LP (CPAL.125.1, CPAL.125.2, CPAL.135, CPAL.144, CPAL.153, CPAL.177, CPAL.181, and CPAL.183), Si/MC (CPAL.146, CPAL.149, CPAL.156, CPAL.172, CPAL.199, CPAL.215, CPAL.219, and MACH.22).
Repository. CPAL-UAEM (19), MACH (1).
Description. Twenty one flat–backed millipedes, head always without eyes, but with Tömösváry organ at the base of antennae, with seven antennomers plus one apical disk; each body ring contains tergites, pleurites, and sternites fused without suture lines, each ring with an anterior prozonite and a laterally projecting metazonite (paranota); CPAL.137 with 17 body rings, CPAL.139 with 19 rings, CPAL.149 with 20 rings, CPAL.153 with 19 rings, CPAL.172 with 12 rings, CPAL.177 with 16 rings, CPAL.178 and CPAL.181 with 17 rings, CPAL.183 with 19 rings, CPAL.215 with 19 rings, CPAL.219 with 20 rings, and MACH.22 with 15 rings.
Comments. The fossil record of Polydesmida is predominantly found in Mexican and Dominican amber (Álvarez-Rodríguez et al., 2024). The Polydesmida fossil record in Mexico contains six families, three genera, and three species. Mexican amber has more families and species of polydesmid millipedes, unlike Dominican amber, which has more reported genera. Compared to other millipede orders in the fossil record, Polydesmida exhibits a larger number of genera.
Suborder Leptodesmidea Brölemann, 1916
Superfamily Chelodesmoidea Cook, 1895
Family Chelodesmidae Cook, 1895
Chelodesmidae indet.
Referred material. one specimen, CPAL.211: ♀.
Locality: Si/MC.
Repository: CPAL-UAEM (1).
Description: Head wider than collum, antennae long, trunk with 17 rings, narrow, broadest at rings 3–5, tapering gradually toward telson; integument smooth and shiny, with a typical pore shape; telson long and conical, flattened dorsoventrally; legs long and slender.
Comments. The family Chelodesmidae is the most diverse group within the order Polydesmida, consisting of 1,437 genera and 3,500 currently living species (Sierwald and Bond, 2007). Its distribution ranges from southern Mexico and the Antilles to northern Argentina and southern Brazil, except Chile (Shelley and Golovatch, 2011; Enghoff, 2015). Fossils of Chelodesmidae have been found in amber from Mexico and the Dominican Republic. †Maatidesmus paachtun Riquelme and Hernández-Patricio, 2014 , has been described in Mexican amber (Riquelme et al., 2014b).
Superfamily Sphaeriodesmoidea Humbert and DeSaussure, 1869
Family Sphaeriodesmidae Humbert and DeSaussure, 1869
Sphaeriodesmidae indet.
Referred material. five specimens, CPAL.130, CPAL.131, CPAL.152, CPAL.184, and CPAL.206.
Locality. Si/CLG (CPAL.152), Si/GV (CPAL.130, and CPAL.131), Si/MC (CPAL.184), Pa/EB (CPAL.206).
Repository. CPAL-UAEM (5).
Description. Head projecting downward, antennae long, dorsally smooth surface, body modified to rotate into a tight ball, CPAL.130 with 20 body rings, CPAL.131 with 14 rings, CPAL.184 with 13 rings, and CPAL.206 with 14 rings, all have narrow sternites, with prozones reduced, tergite 3 and 4 enlarged, telson flattened dorsoventrally, and legs slender.
Comments. This Mexican record represents the oldest known fossil of the family Sphaeriodesmidae. Previously, †Cyclodesmus porcellanus Pocock, 1894 was described from the Late Pleistocene of Jamaica (Baalbergem and Donovan, 2012). Sphaeriodesmidae consists of 14 genera and 91 extant species, which are found in Central America and the Greater and Lesser Antilles, as well as from the northern to the southern United States (Shelley and Golovatch, 2011; Enghoff, 2015).
Suborder Polydesmidea Pocock, 1887
Infraorder Oniscodesmoides Simonsen, 1990
Superfamily Pyrgodesmoidea Silvestri, 1896
Family Pyrgodesmidae Silvestri, 1896
Pyrgodesmidae indet. (Figure 2a-b)
Referred material: 19 specimens, CPAL.120 (Figure 2a), CPAL.128, CPAL.136, CPAL.138, CPAL.155: ♀, CPAL.158, CPAL.167: ♀, CPAL.168, CPAL.173: ♀, CPAL.176: ♀, CPAL.185: ♀, CPAL.186, CPAL.187: ♀, CPAL.200, CPAL.205: ♀, CPAL.210: ♀, CPAL.214: ♀ (Figure 2b), CPAL.217, new record CAF-6.
Locality: Si/EC (CPAL.167, CPAL.168, and CPAL.187), Si/EPo (CPAL.128), Si/GV (CPAL.120, and CPAL.138), Si/LP (CPAL.176, CPAL.185, CPAL.186, CPAL.200, and CPAL.205), Si/MC (CPAL.136, CPAL.155, CPAL.158, CPAL.173, CPAL.214, CPAL.217, new record CAF-5), To/RS (CPAL.210).
Repository. CPAL-UAEM (18), new record CAF (1).
Description. Head small, granular, projecting downward, entirely covered by the collum. Collum with aligned frontal lobes; small, wide trunk, surrounded by long rows of tubercles; porosteles present (Hoffman et al., 1996). CPAL.167, with 18 rings; CPAL.168, divided into two parts, anterior head + 10 rings; CPAL.173 and CPAL.176, with 19 rings; CPAL.185, with 20 rings; CPAL.186, with 10 rings; CPAL.187, with 18 rings; CPAL.214 and CPAL.217, with 18 rings; CPAL.205 and CPAL.210, with 17 rings.
Comments: The family Pyrgodesmidae is found in tropical regions, extending from southern Europe and North Africa to Japan, Taiwan, central China, and the southern United States (Hoffman et al., 1996). Pyrgodesmidae includes 140 genera and around 400 living species (Enghoff, 2015). †Myrmecodesmus antiquus Riquelme and Hernández-Patricio, 2021 has been described in Mexican amber (Riquelme et al., 2021).
Infraorder Polydesmoides Pocock, 1887
Superfamily Trichopolydesmoidea Verhoeff, 1910
Family Trichopolydesmidae Verhoeff, 1910
Trichopolydesmidae indet.
Referred material. three specimens, CPAL.119: ♀, CPAL.121: ♂, and CPAL.150: ♂.
Locality: Si/GV(CPAL.121), Si/LP (CPAL.119, and CPAL.150).
Repository. CPAL-UAEM (3).
Description. Small polydesmid millipedes, head with slightly granular surface with scattered short setae in a line from the vertex to the clypeus, with slender antennae. Trunk short, CPAL.121 with 20 rings and CPAL.150 with 18 rings, paraterga lobed, raised laterally, lacking transverse suture, dorsal surface granulated, with rows of setae; ozopores present on both sides, with a typical arrangement; legs covered with fine setae.
Comments. The family Trichopolydesmidae comprises 75 genera and 140 species, primarily found in the northern hemisphere and temperate and tropical regions (Enghoff et al., 2015). There were no reports of fossil records prior to this study. This represents the earliest fossil record from the Oligo-Miocene strata in the tropical regions of North America.
Superorder Nematophora Verhoeff, 1913
Order Stemmiulida Cook, 1895
Family Stemmiulidae Pocock, 1894
Stemmiulidae indet.
Referred material. one specimen, CPAL.141: ♀.
Locality. Si/LP.
Repository. CPAL-UAEM (1).
Description. Head with ocelli on each side, long and thin antennae, eight antennomeres, and a terminal apical disc; collum larger than the following first tergites; trunk long, cylindrical, with 36 rings, pleurotergites with a broad dorsal suture, unfractionated prozonites and metazonites, covered by fine striations, ozopores on the fifth segment; legs long with eight podomeros; telson short, with spineteres on the posterior edge of the epiproct.
Comments. The fossil record of Stemmiulida is represented by amber inclusions from the lowermost Late Cretaceous of Myanmar, the Oligo-Miocene of Mexico, and the Early Miocene of the Dominican Republic (Álvarez-Rodríguez et al., 2024). †Parastemmiulus elektron Riquelme et al., 2013 has been previously described in Mexican amber.
Superorder Juliformia Attems, 1926
Order Julida Brandt, 1833
Julida indet.
Referred material. one specimen, CPAL.124: ♀.
Locality. Si/GV.
Repository. CPAL-UAEM (1).
Description. CPAL.124 has a long, uniform body that is vermiform in shape. Head lacks a median suture extending to the labrum, eyes oval, with two long frontal setae; trunk with 38 rings; tergites and pleurites fused in a cylindrical shape, dorsally striated; it is identified as female, with no modification to the first pair of legs.
Comments. Julida is found across a wide Holarctic distribution, spanning from southern North America to Guatemala and extending throughout most of Eurasia, including Japan and Indochina (Shelley and Golovatch, 2011). This order contains 16 families, 18 genera, and 739 species (Shear, 2011). In contrast, the fossil record of Julida comprises two families, three genera, and 11 species (Álvarez-Rodríguez et al., 2024).
Unassigned Diplopoda
Referred material. (8 specimens) CPAL.151, CPAL.154, CPAL.159, CPAL.170, CPAL.171, CPAL.179, CPAL.208, and CPAL.216.
Locality. Si/CLG (CPAL.151), Si/EC (CPAL.159), Si/Pa (CPAL.179), Si/LP (CPAL.154), Si/MC (CPAL.208, and CPAL.216), Hu/ER (CPAL.170, and CPAL.171).
Repository. CPAL-UAEM (8).
Description. Recognizable millipede forms, head differentiated, no ocelli, antennae with seven antennomers plus one apical disk. CPAL.154 with 18 body rings; CPAL 159 with indeterminate fragmented rings; CPAL.170 and CPAL.171 juvenile of indeterminate family; CPAL.179 without posterior rings or telson; CPAL.208 with 12 posterior rings, neither the head nor the anterior rings were preserved.
Comments. Due to their incomplete condition, characterized by fragmented or headless bodies, or because they are juvenile specimens lacking recognizable diagnostic features, these fossils are not categorized at the specific order or family level. However, they do represent distinctive fossil forms within Diplopoda.
5. Conclusions
The present checklist of Mexican amber millipedes complements the global catalog of fossil Diplopoda compiled by Álvarez-Rodríguez et al. (2024). It includes descriptions and comments on each record excluded from the worldwide catalog due to its format limitations, as well as with new findings.
As a result of the previous compilations by Riquelme and Hernández-Patricio (2018), Álvarez-Rodríguez et al. (2024), and this current study, the Mexican fossil record now includes a total of 120 amber inclusions. These inclusions are classified into seven orders, eight families, five genera, and four species. The recognized orders include Siphonophorida, Spirobolida, Stemmiulida, Polydesmida, Polyxenida, Platydesmida, and Julida. Polydesmida is the majority, representing 66% of the total fossil inclusions. Family-level diversity includes the families Siphonophoridae, Stemmiulidae, Xystodesmidae, Platyrhacidae, Chelodesmidae, Pyrgodesmidae, Sphaeriodesmidae, and Trichopolydesmidae, as well as the genera Siphonophora, Parastemmiulus, Anbarrhacus, Maatidesmus, and Myrmecodesmus; and the species †Parastemmiulus elektron, †Anbarrhacus adamantis, †Maatidesmus paachtun, and †Myrmecodesmus antiquus.
Additionally, this work expands the identification of new records for the families Chelodesmidae (1 record), Pyrgodesmidae (19 records), Sphaeriodesmidae (5 records), and Trichopolydesmidae (3 records). However, due to conservation constraints, over 20 fossil inclusions have been provisionally identified only at the order level.
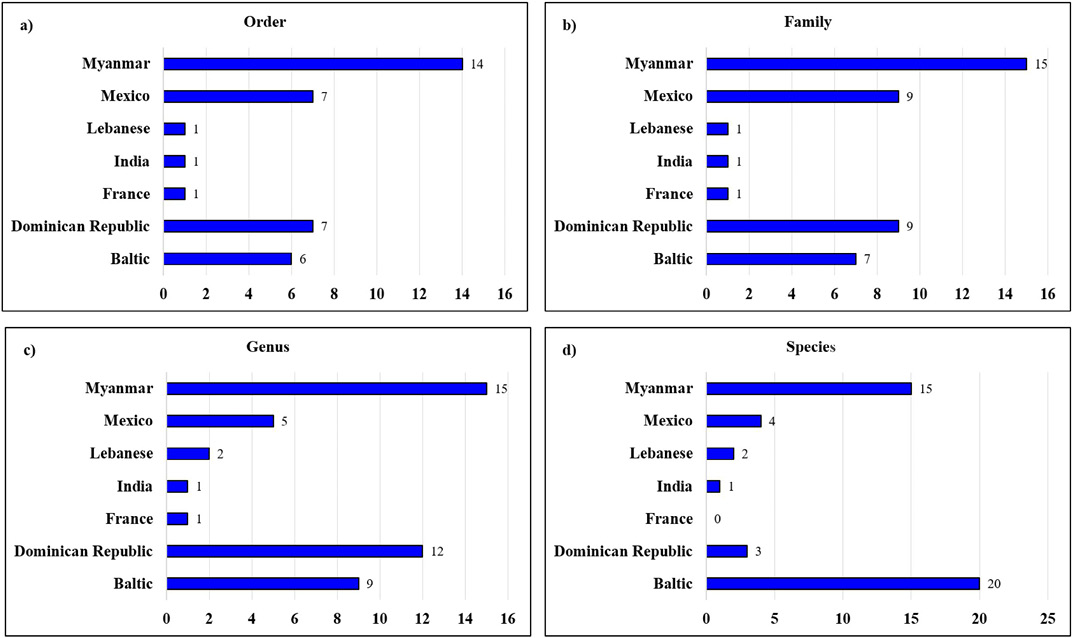
The Mesozoic fossil record of millipedes (Álvarez-Rodríguez et al., 2024), mainly found in amber deposits from the lowermost Late Cretaceous of Myanmar, indicates a rise in diversity at higher taxonomic ranks, such as orders and families (Figure 3). However, this diversity declined during the Cenozoic, as shown by the fossil records from Baltic amber, and more recent deposits from Mexico and the Dominican Republic (Figure 3). According to our results, the fossil record from Mexico show a reduced level of variation at both the order and family levels compared to that observed in Myanmar amber.
Finally, checklists are structured tools to verify biological data according to established nomenclature and taxonomic frameworks. They are essential resources for research and conservation efforts focused on biodiversity. In this context, the current work, which incorporates referred material deposited in established collections, highlights the fossil diversity of Diplopoda in southern North America and supports initiatives dedicated to the preservation of paleontological resources in Mexico (Riquelme and Hernández-Patricio, 2018).
Acknowledgments
We thank Luis Zúñiga for access to the MALM collection, Bibiano Luna for the MACH collection, and Victor Córdova for the CAF-IPN collection. The fieldwork was conducted with the invaluable assistance of Patricia Díaz, who supports indigenous mining and artisanal communities in Simojovel. We thank Susana Guzmán-Gómez at the LMF2-LANABIO, Instituto de Biología-UNAM, for photomicrography assistance. We also appreciate the feedback from Editor-in-Chief Josep A. Moreno Bedmar and two anonymous reviewers, which helped improve the paper after publication. We thank Sandra Ramos, León Felipe Alvarez, and the editorial team for their help with editing and proofreading. The SECIHTI grant supported MAR as part of the MMRN postgraduate program at the UAEM.
References
Allison, R. C. (1967). The Cenozoic Stratigraphy of Chiapas, Mexico, with Discussions of the Classification of the Turritellidae and Selected Mexican representatives. [Unpublished PhD Tesis], University of California at Berkley.
Álvarez-Rodríguez, M., Riquelme, F., Hernández-Patricio, M., & Cupul-Magaña, F. (2024). Diplopoda in the world fossil record. Zoological Systematics, 49(3), 185–245. https://doi.org/10.11865/zs.2024201
Baalbergen, E., & Donovan, S. K. (2012). Terrestrial arthropods from the Upper Pleistocene of Jamaica: systematics, palaeoecology and taphonomy. Geological Journal, 48(6), 628–645. http://dx.doi.org/10.1002/gj.2477
Bueno-Villegas J., Sierwald, P., & Bond J. E. (2004). Diplopoda. In B. Llorente, J. Morrone, O. Yañez, & I. Vargas (Eds.), Biodiversidad, Taxonomía y Biogeografía de Artrópodos de México. Hacia una síntesis de su conocimiento (4th, pp. 569–599). UNAM-CONABIO, México. https://doi.org/10.21829/azm.2014.303100
Edgecombe, G. D. (2015). Diplopoda: Fossils. In A. Minelli (Ed.), Treatise on Zoology Anatomy, Taxonomy, Biology. The Myriapoda, Vol.2 (pp. 337–352). Brill, Netherlands. https://doi.org/10.1163/9789004188273
Enghoff, H. (2015). Diplopoda: Geographical distribution. In A. Minelli (Ed.), Treatise on Zoology Anatomy, Taxonomy, Biology. The Myriapoda, Vol. 2 (pp. 329–336). Brill, Netherlands. https://doi.org/10.1163/9789004188273
Enghoff, H., Golovatch, S., Short, M., Stoev, P., & Wesener, T. (2015). Diplopoda: Taxonomic overview. In A. Minelli (Ed.), Treatise on Zoology Anatomy, Taxonomy, Biology. The Myriapoda, Vol. 2 (pp. 363–453). Brill, Netherlands. https://doi.org/10.1163/9789004188273
Frost, S. H., & Langenheim, R. L. (1974). Cenozoic Reef Biofacies, Tertiary Larger Foraminifera and Scleractinian Corals from Chiapas, Mexico. Northern Illinois University Press, De Kalb.
Graham, A. (1999). Studies in neotropical paleobotany. XIII. An Oligo-Miocene palynoflora from Simojovel (Chiapas, Mexico). American Journal of Botany, 86, 17–31. https://doi.org/10.2307/2656951
Golovatch, S. I. (2013). A reclassification of the millipede superfamily Trichopolydesmoidea, with descriptions of two new species from the Aegean region (Diplopoda, Polydesmida). ZooKeys, 340, 63–78. https://doi.org/10.3897/zookeys.340.6295
Hoffman, R. L., Golovatch, S. I., Adis, J., & Demarais, J. W. (1996). Practical keys to the orders and families of millipedes of the Neotropical region (Myriapoda: Diplopoda). Amazoniana, 14, 1–35.
Hoffman, R. L. 1999. Checklist of the millipeds of north and middle America. (Special publication no. 8), Virginia Museum of Natural History. https://www.biodiversitylibrary.org/item/335228
Koch, M. (2015). Diplopoda: General morphology. In A. Minelli (Ed.), Treatise on Zoology Anatomy, Taxonomy, Biology. The Myriapoda (2nd, pp. 7–69). Brill, Netherlands. https://doi.org/10.1163/9789004188273
Langenheim, J. H. (1966). Botanical source of amber from Chiapas, Mexico (Fuente botánica del ámbar de Chiapas, México). Ciencia, 24(5–6), 201–210.
Langenheim, J. H. (2003). Plant Resins: Chemistry, Evolution, Ecology, and Ethnobotany. Timber Pres. https://doi.org/10.1600/036364404772974374
Latreille, P. A. (1802). Histoire naturelle, générale et particulière des Crustacés et des Insectes. Dufart.
Lucas, H. (1840). Histoire Naturelle des Crustacés, des Arachnides et des Myriapodes. Duménil.
Perrilliat, M., Vega, F., & Coutiño, M. (2010). Miocene mollusks from the Simojovel area in Chiapas, southwestern Mexico. Journal of South American Earth Sciences, 30(2), 111–119. https://doi.org/10.1016/j.jsames.2010.04.005
Riquelme, F., & Hernández-Patricio, F. (2018). The millipedes and centipedes of Chiapas amber. Check List, 14, 637–646. https://doi.org/10.15560/14.4.637
Riquelme, F., Alvarado-Ortega, J., Ramos-Arias, M., Hernández, M., le Dez, I., Lee-Whiting, T. A., & Ruvalcaba-Sil, J. L. (2013). A fossil stemmiulid millipede (Diplopoda: Stemmiulida) from the Miocene amber of Simojovel, Chiapas, Mexico. Historical Biology, 26(4), 415–427. https://doi.org/10.1080/08912963.2013.778843
Riquelme, F., Ruvalcaba-Sil, J. L., Alvarado-Ortega, J., Estrada-Ruiz, E., Galicia-Chávez, M., Porras-Múzquiz, H., Stojanoff, V., Siddons, D. P., & Miller, L. (2014a). Amber from México: Coahuilite, Simojovelite and Bacalite. MRS Proceedings, 1618, 169–180. https://doi:10.1557/opl.2014.466
Riquelme, F., Hernández-Patricio, M., Martínez-Dávalos, A., Rodríguez-Villafuerte, M., Montejo-Cruz, M., Alvarado-Ortega, J., Ruvalcaba-Sil, J., & Zúñiga-Mijangos, L. (2014b). Two flat-backed polydesmidan millipedes from the Miocene Chiapas-Amber Lagerstätte, Mexico. PLoS ONE, 9, e105877. https://doi.org/10.1371/journal.pone.0105877
Riquelme, F., Hernández-Patricio, M., & Álvarez-Rodríguez, M. (2021). A Miocene pyrgodesmid millipede (Polydesmida: Pyrgodesmidae) from Mexico. PeerJ, 9(2), e10574. http://dx.doi.org/10.7717/peerj.10574
Riquelme, M., Ortega-Flores, B., Estrada-Ruiz, E., & Córdova-Tabares, V. (2025). Zircon U–Pb ages of the Chiapas Amber-Lagerstätte in the uppermost Simojovel Formation, southwest Mexico. Canadian Journal of Earth Sciences, 62(6), 1–16. https://doi.org/10.1139/cjes-2024-0100
Shear, W. A. (2011). Class Diplopoda de Blainville in Gervais, 1844. In Z. Q. Zhang (Ed.), Animal Biodiversity: An Outline of Higher-level Classification and Survey of Taxonomic Richness. (3148, pp. 169–164.) Zootaxa. https://doi.org/10.11646/zootaxa.3148.1.32
Shelley, R. M., & Golovatch, S. I. (2011). Atlas of Myriapod Biogeography. I. Indigenous Ordinal and Supra-Ordinal Distributions in the Diplopoda: Perspectives on Taxon Origins and Ages, and a Hypothesis on the Origin and Early Evolution of the Class. Insecta Mundi, 158, 1–134.
Sierwald, P., & Bond, J. (2007). Current Status of the Myriapod Class Diplopoda (Millipedes): Taxonomic Diversity and Phylogeny. Annual Review Entomology, 52, 401–20. https://doi.org/10.1146/annurev.ento.52.111805.090210
Sierwald, P., & Spelda, J. (2019). MilliBase. http://www.millibase.org
Wesener, T., & Moritz, L. (2018). Checklist of the Myriapoda in Cretaceous Burmese amber and a correction of the Myriapoda identified by Zhang (2017). Check List, 14(6), 1131–1140. http://dx.doi.org/10.15560/14.6.1131.