Ankylosaurians from Coahuila, Mexico
Anquilosaurios de Coahuila, México
Rivera-Sylva, Héctor E.1,* ; Aguillón-Martínez, Martha C.1;
; Aguillón-Martínez, Martha C.1;
Guzmán-Gutiérrez, José Rubén2; Flores-Ventura, José1
1 Museo del Desierto, Carlos Avedrop Dávila 3745, 25022, Saltillo, Coahuila, Mexico.
2 Universidad Humanista de las Américas, Martín de Zavala 510, 64000, Monterrey, Nuevo León, México.
* hrivera@museodeldesierto.org
Abstract
The fossil record of Ankylosauria is well-documented in several regions of the world yet remains relatively scarce in Mexico. Here we report new thyreophoran material from the Late Cretaceous in Coahuila, northeastern Mexico. The specimens, which include isolated osteoderms and partial skeletal elements, contribute to our understanding of the diversity, paleobiogeography, and paleoecology of this dinosaur clade in North America. Morphological comparisons suggest affinities with nodosaurid ankylosaurs, expanding their known range and ecological roles in the region. This discovery highlights the importance of Coahuila as a key locality for studying dinosaur faunas at the southern margin of the Western Interior Seaway.
Keywords: Ankylosauria, Ankylosauridae, Coahuila, diversity, Nodosauridae.
Resumen
El registro fósil de Ankylosauria está bien documentado en varias regiones del mundo, pero sigue siendo relativamente escaso en México. En este trabajo reportamos nuevo material de tireóforos del Cretácico Tardío en Coahuila, noreste de México. Los especímenes, que incluyen osteodermos aislados y elementos esqueléticos parciales, contribuyen a nuestra comprensión de la diversidad, paleobiogeografía y paleoecología de este clado de dinosaurios en América del Norte. Las comparaciones morfológicas sugieren afinidades con anquilosaurios nodosáuridos, ampliando su rango conocido y sus roles ecológicos en la región. Este descubrimiento resalta la importancia de Coahuila como una localidad clave para el estudio de faunas de dinosaurios en el margen sur del Mar Interior Occidental.
Palabras clave: Ankylosauria, Ankylosauridae, Coahuila, diversidad, Nodosauridae.
1. Introduction
The monophyletic Ankylosauria (Osborn, 1923) is a clade within Thyreophora which includes Ankylosauridae and Nodosauridae and all taxa more closely related to these dinosaurs than to Stegosaurus (Carpenter, 1990), forming together the clade Thyreophora (Nopcsa, 1915), which includes ankylosaurs and stegosaurs and all taxa more closely related to them than to Cerapoda (Ornithopoda + Marginocephalia). They were a very diverse group of thyreophoran dinosaurs characterized by flat and wide skulls with leaf-like teeth, a heavily armored of the skull and body protected by a shield of densely packed osteoderms, a barrel-shaped body, and an obligatory quadrupedal stance (Vickaryous et al., 2004; Carpenter, 2012).
There are two formally distinguished families: Ankylosauridae, and Nodosauridae (Coombs, 1978; Arbour and Currie, 2016). They differ by a number of diagnostic characters identified (e.g. in the skull, and teeth), but they are most easily distinguished by the presence (Ankylosauridae) or absence (Nodosauridae) of a distinctive tail club formed by the co-ossification of the caudal-most vertebrae combined with a series of plate-like osteoderms.
The earliest valid members of Ankylosauridae are known from the Middle Jurassic of England and China (Carpenter, 2012). Ankylosaurs have been found on every continent including Africa. The highest diversity of this group is observed during the Late Cretaceous in Asia, North America and Europe. Ankylosaurs are also reported, however to a much lesser degree of diversity from South America, and Antarctica (Arbour and Currie, 2016).
Ankylosaurs are characteristic and abundant components of dinosaur assemblages from the Late Cretaceous of western North America (Laramidia). Campanian strata yield numerous specimens of nodosaurids and ankylosaurids, mostly from the north of the continent. In contrast, the record from the southern part of Laramidia (e.g., Mexico) is surprisingly scarce (Loewen et al., 2013; Rivera-Sylva and Carpenter, 2014).
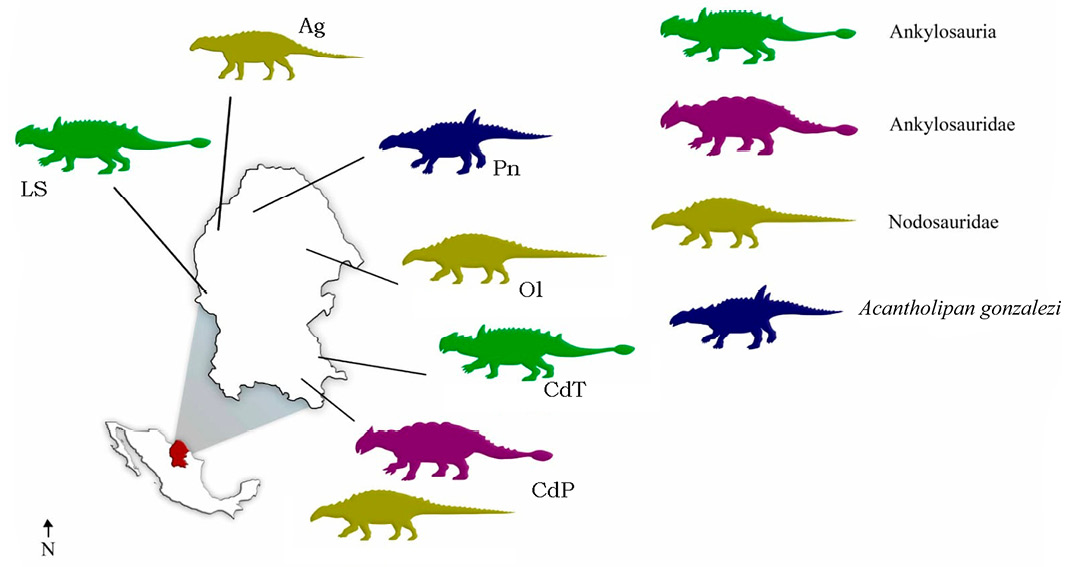
Ankylosaur specimens are extremely rare in Upper Cretaceous deposits of Mexico, which contrasts with the rich hadrosaurian assemblages reported from the region (Rivera-Sylva and Carpenter, 2014). Nevertheless, ankylosaur remains have repeatedly been reported from Mexican strata, although most of these are undiagnostic consisting of a few teeth, osteoderms, vertebrae, and long bones (Rivera-Sylva et al., 2011a; Figure 1). To date, Acantholipan gonzalezi is the only thyreophoran formally described for Mexico (Rivera-Sylva et al., 2018). The material of ankylosaur remains documented so far, comes from the Zorrillo-Tabernas Indiferenciadas Formation (Bajocian) of Puebla, the Mojado Formation (latest Albian-earliest Cenomanian) of Chihuahua (which is the first report for Mexico of Ankylosaurian tracks), El Gallo Formation (Campanian) of Baja California, the San Carlos Formation (Campanian) of Chihuahua, the Cerro del Pueblo Formation (Campanian) and the Olmos Formation (Campanian-Maastrichtian) of Coahuila (Morris, 1967; Rivera-Sylva and Espinosa-Chávez, 2006; Kappus et al., 2011; Martínez-Díaz and Montellano-Ballesteros, 2011; Rivera-Sylva et al., 2011a; Rodríguez-de la Rosa et al., 2018; Rivera-Sylva et al., 2019; Rivera-Sylva et al., 2021).
An ankylosaurid femur from the Olmos Formation near Sabinas, Coahuila, was mentioned by Meyer et al. (2005), but after a close examination of the specimen we conclude that it does not belong to Ankylosauria but rather represents an unidentified ornithopod.
This paper describes newly discovered material that sheds light on the diversity of ankylosaurs in Mexico, offering a fresh taxonomic perspective. These findings are particularly significant given the scarcity of ankylosaur remains in the region, which has historically limited our understanding of their distribution and diversity. The new material not only provides insights into their morphological variability but also underscores the importance of Mexico as a region for studying ankylosaurs and their broader paleobiogeographic implications.
Abbreviations: CPC = Colección Paleontológica de Coahuila, Museo del Desierto, Mexico; MUDE = Museo del Desierto, Mexico.
2. Material and methods
The material described herein from the southeast of Coahuila was discovered during field seasons between 2000 and 2002. All the material is housed in the Colección Paleontológica de Coahuila at the Museo del Desierto in Saltillo, Coahuila. The exact locality information of each specimen is on file at the Museo del Desierto, Saltillo, Coahuila.
The specimens were measured using a digital caliper, and photographed using a Canon EOS Rebel T2i with a Canon Zoom Lens EF 35-80mm 1:4-5.6 III with filters close ups 1-3, and a Nikon COOLPIX P610 with a lens NIKKOR 60X Wide Optical Zoom ED VR (4.3–258 mm 1:3.3–6.5).
3. Systematic palaeontology
Dinosauria Owen, 1842
Ornithischia Seeley, 1888
Thyreophora Nopcsa, 1915
Eurypoda Sereno, 1986
Ankylosauria Osborn, 1923
Ankylosauria indet.
Material. Caudal vertebra (CPC 3045; Figure 2 a–d).
Horizon and locality. Soledad Beds (Campanian-Maastrichtian, Late Cretaceous), 22 km southwest of the town La Esmeralda; municipality of Sierra Mojada, Coahuila, Mexico.
Description. CPC 3045 is a badly weathered centrum with a hexagonal cross-section, and pronounced transverse processes, the latter being completely fused with the mid part of the neurocentral junction. The neural canal is circular in cross-section. Measurements of CPC 3045 in millimetres: length = 70; maximum height = 60; maximum width = 98.
Discussion. The well-developed transverse process suggest that the specimen represents an anterior caudal vertebra of an ankylosaurian (Vickaryous et al., 2004; Arbour, 2009). The specimen was mention as an hadrosaur caudal by Rivera-Sylva and Carpenter (2014).
Material. Caudal vertebra (CPC 3046; Figure 2 e–g).
Horizon and locality. Aguja Formation (Campanian, Late Cretaceous), La Salada locality; municipality of Ocampo, Coahuila, Mexico.
Description. CPC 3046 is a badly weathered amphicoelus centrum of a proximal caudal vertebra, with a rounded cross-section, and pronounced transverse processes, the latter being completely fused with the mid part of the neurocentral junction. The neural canal is circular in cross-section. Measurements of CPC 3046 in millimetres: length = 88; maximum height = 178; maximum width = 240.
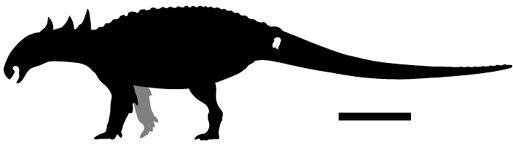
Discussion. The well-developed transverse process and the rounded outline suggest that the specimen represents an anteriormost caudal vertebra of an ankylosaurian (Vickaryous et al., 2004; Arbour, 2009). The specimen's size aligns with that of a significantly large ankylosaurid, estimated at approximately 7 meters in length, as determined through comparisons with the caudal vertebrae of other ankylosaurians (e.g., Peloroplites cedrimontanus) (Carpenter, pers. comm., 2018; Molina-Pérez, pers. comm., 2023; Figure 3).
In 2017, Shiller described a right tibia discovered in the Aguja Formation of northern Coahuila, which is part of a private collection. Consequently, the precision of the measurement could not be verified. Nevertheless, if the measurement is accurate, it may pertain to a significantly large specimen (Molina-Pérez, pers. comm., 2023), consistent with the attributes of CPC 3046.
Material. Caudal vertebrae (CPC 3047, a, b, c; Figure 2 h–p).
Horizon and locality. Cerro del Pueblo Formation (Upper Cretaceous, Campanian), Rincón Colorado locality, General Cepeda, Coahuila, Mexico.
Description. These are three amphicoelus centra of caudal vertebrae. CPC 3047 had a centrum with circular cross-section being as long as is tall, while CPC 3047b and c had centra with a hexagonal cross-section, with its length been half of its height. All three vertebrae had broad bases of transverse processes with oval cross-section, the latter being fused with the mid part of the neurocentral junction. Measurements of CPC 3047a in millimetres: length = 46.1; maximum height = 58.6; maximum width = 72. CPC 3047b: length = 36.9; maximum height = 58.6; maximum width = 65.9. CPC 3047c: length = 37.8; maximum height = 54.4; maximum width = 64.7.
Discussion. The well-developed transverse process and the rounded centrum of CPC 3047a suggest that the specimen represents an anterior caudal vertebra of an ankylosaurian; while CPC 3047 b and c are mid-caudals (Vickaryous et al., 2004; Arbour et al., 2009).
Material. Right metatarsal II (CPC 2975; Figure 4).
Horizon and locality. The specimen come from the Cerro del Pueblo Formation (Upper Cretaceous, Campanian), Porvenir de Jalpa locality, General Cepeda, Coahuila, Mexico. Description. The proximal articulation of the metatarsal is rugose and bulbous, subrectangular, dorsally inclined at an angle of approximately 10°, obliquely oriented relative to the long axis of the shaft, being in consequence the ventral edge longer than the dorsal, and is dorsally smooth, although some parts are weathered. The rugose texture extends distally on the medial side in a ridge to the mid-length of the shaft. The distal condyle extends proximally onto the plantar surface on the medial and lateral sides. Measurements of CPC 2975 in millimetres: length = 103.5; maximum height = 41.1; maximum width = 68.7.
Discussion. The right metatarsal II CPC 2975 is as described and figure of those of Niobrarasaurus coleii, Europelta carbonensis, and for the ankylosaur AMNH 5266 (Carpenter et al., 1995; Coombs, 1986; Kirtkland et al., 2013), therefore is unequivocal ankylosaurian.
Material. Phalanges (CPC 3048a, b; Figure 5 p–u).
Horizon and locality. Cañón del Tule Formation (Upper Cretaceous, Maastrichtian), Carolinas locality, Ramos Arizpe, Coahuila, Mexico.
Description. They are anteroposteriorly short. With a shallow articular surface and poorly developed distal condyles. Measurements of CPC 3048a in millimetres: length = 32.4; width = 43.7; maximum height = 21.5. CPC 3048b in millimetres: length = 25; width = 35.8; maximum height = 16.2.
Discussion. The phalanges are similar of those described for Peloroplites cedrimontanus (Carpenter et al., 2008).
Material. Unguals (CPC 3049; CPC 3050; CPC 3051; Figure 5 a–l).
Horizon and Locality. Cerro del Pueblo Formation (Upper Cretaceous, Campanian), Rincón Colorado locality, General Cepeda, Coahuila, Mexico.
Description. The three unguals are widest at their proximal articulation and taper distally to a blunt tip. CPC 3049 and CPC 3050 are as long as there are wide; CPC 3051 is incomplete but is longer than wide. They are rugose both dorsally and ventrally, and on the ventral surface the neck is tucked, and bounded on either side by a proximally extending horn. Measurements of CPC 3049 in millimetres: length = 64.7; width = 52.5; maximum height = 22.5. CPC 3050 in millimetres: length = 61.7; width = 53.8; maximum height = 69.7. CPC 3051 in millimetres: length = 52.5; width = 39; maximum height = 18.
Discussion. These unguals are blunt as seen in other ankylosaurians (Carpenter, 1990; Carpenter et al., 1995; Arbour et al., 2009; Currie et al., 2011) rather than hoofed liked as seen in ceratopsians.
Material. Ungual (CPC 3052; Figure 5 m–o).
Horizon and Locality. Cañón del Tule Formation (Upper Cretaceous, Maastrichtian), Carolinas locality, Ramos Arizpe, Coahuila, Mexico.
Description. The ungual is widest at their proximal articulation and taper distally to a blunt tip, being as long as it is wide; with a rugose texture dorsally and ventrally. Measurements of CPC 3052 in millimetres: length = 60.6; width = 57.7; maximum height = 22.6.
Discussion. The ungual is blunt as seen in other ankylosaurians (Carpenter, 1990; Carpenter et al., 1995; Arbour et al., 2009; Currie et al., 2011).
Ankylosauridae Brown, 1908
Ankylosauridae indet.
Material. CPC 3053a, ungual; CPC 3053b, osteoderm; (Figure 6 e–g).
Horizon and locality. Cerro del Pueblo Formation (Upper Cretaceous, Campanian), La Parrita locality, General Cepeda, Coahuila, Mexico.
Description. The ungual (CPC 3053a) is broken and badly weathered, the tip is preserved and the left side. In the right side can be see the proximal end of the vascular groove, the dorsal surface is rugose while the ventral is smoother with some foramina present. The osteoderm (CPC 3053b) is with only right side preserved, with a dorsal surface with a rise keel orientated obliquely, with a fine pitting in its dorsal surface, with a convex ventral surface. Measurements in millimetres of CPC 3053a: length = 66.1; maximum height = 26.2; maximum width = 54.9. CPC 3053b: length = 47.6; maximum height = 24.1; maximum width = 48.1.
Discussion. The ungual is as seen in other ankylosaurians (Carpenter, 1990; Carpenter et al., 1995; Arbour et al., 2009; Currie et al., 2011). The basally-excavated ostederm is ankylosaurian-type as seen in other Ankylosauridae as Ahshislepelta minor (Burns and Sullivan, 2011).
Material. CPC 3054, osteoderm; (Fig. 6 a–d).
Horizon and Locality. Cerro del Pueblo Formation (Upper Cretaceous, Campanian), Rancho Ammonite locality, General Cepeda, Coahuila, Mexico.
Description. The osteoderm (CPC 3054) has a dorsal surface with a rise blunt keel orientated caudally, and with convex ventral surface. It has a smooth surface with vascular channels over its surface. Is thick-walled and thicker along the keel. The base is oval, longer than wide, although only its left side is the only preserved edge. Measurements in millimetres of CPC 3054: length = 33.9; maximum height = 14; maximum width = 24.9.
Discussion. The concave ventral surface indicates ankylosauridae affinity (Coombs & Maryańska, 1990), although its small size cannot be determined if belong to an immature individual or was a small ossicle in a large individual.
Nodosauridae Marsh, 1890
Acantholipan gonzalezi Rivera-Sylva et al., 2018
The only thyreophoran formally described for Mexico is the nodosaurid Acantholipan gonzalezi (Rivera-Sylva et al., 2018). It is a nodosaur that inhabited on the northwest coast of Coahuila, which today is a semi-desert around the township of Ocampo. The sediments in this region correspond to the Pen Formation, with an age of about 85 million years. According to the remains preserved, the individual was a juvenile with a length of about 4.5 meters and a mass close to 500 kg. Its remains were extracted from a bedrock that was deposited in shallow coastal seawaters. The carcass floated down a river prior to deposition, a taphonomical scenario frequently observed among nodosaurs (e.g., Aletopelta, Borealopelta, Nodosaurus, Stegopelta, Pawpawsaurus, and Niobrarasaurus; Carpenter et al., 1995; Carpenter and Kirkland, 1998; Ford and Kirkland, 2001; Arbour et al., 2016; Brown et al., 2017), unquestionable evidence that the group occupied the ecological niche of nowadays hippopotamus.
The holotype of Acantholipan gonzalezi (CPC 272) consist of a thoracic and a caudal vertebra, the fragment of a rib, the distal end of left humerus, the distal end of the left femur, and a thoracic osteodermal spine. It is distinguished from the other species of nodosaurids due to the following combination of characteristics: ulna with a prominent anterolateral process, tapered and proximally truncated, triangular in lateral view, conical and slightly curved caudally thoracic osteoderm with an oval longitudinal cross-section and vascular grooves on the lateral view.
The spine belongs to the thoracic area based on its circular base and cross section as observed in other nodosaurids (Ford, 2000; Carpenter, 2012; Ősi and Pereda-Suberbiola, 2017), while the spines from the cervical region have a triangular base, the bodies are compressed with a y-shaped cross-section with the dorsal side keeled for most of its length (Ford, 2000; Kirkland et al., 2013; Penkalski, 2014).
The spine rows in Acantholipan and other nodosaurids (e.g., Europelta, Struthiosaurus) might have extended down through the tail, as in extant crocodiles. The spines working with the other body osteoderms may have functioned as an efficient distribution of absorbed radiant heat during basking (both in and out of the water) as in crocodiles (Seidel, 1979; Clarac and Quilhac, 2019). The argument favoring protection is weakened (but not dismissed) when considering the vascularity of osteoderms suggesting an important physiological role in calcium remobilization as suggest by Brum et al. (2023). Therefore, in large semiaquatic reptiles, basking is related to the presence of dermal armor, where the sculpted armor is present only on surfaces that would be available for absorption of solar radiation. These surfaces are exposed to sunlight during basking on land and while the animal is partly submerged (Seidel, 1979).
Nodosauridae indet.
Material. Caudal vertebra; phalanges; osteoderms (CPC 3055; Figure 7).
Horizon and locality. Cerro del Pueblo Formation (Upper Cretaceous, Campanian), La Parrita locality, General Cepeda, Coahuila, Mexico.
Description. The vertebra is slightly amphicoelus. With rounded centra in anterior and posterior view. Is as long as it is tall. The base of the neural canal is rounded. It has a broad base of the transverse process, being on straight angle with the centrum. The phalanges are anteroposteriorly short. With a shallow articular surface and poorly developed distal condyles. CPC 3055b is longer than it is wide, while CPC 3055c is wider than it is long. Measurements of CPC 3055a in millimetres: length = 36.2; maximum height = 39.7; maximum width = 62.9; CPC 3055b in millimetres: length = 55.1; maximum height = 36.5; maximum width = 59.7; CPC 3055c in millimetres: length = 25.7; maximum height = 21.4; maximum width = 43.5. CPC 3055d in millimetres: length = 30; maximum height = 29; maximum width = 22; CPC 3055e in millimetres: length = 25.6; maximum height = 20.3; maximum width = 29.8.
Discussion. The well-developed transverse process and the rounded centrum of CPC 3055a suggest that the specimen represents an anterior caudal vertebra (Vickaryous et al., 2004; Arbour et al., 2009). The phalanges are as those figured by Carpenter (1990). While the fragments of the osteoderm looks flat in their ventral surface, it suggests the nodosaurid affinity of the specimen (Coombs & Maryańska, 1990).
Material. CPC 3092 a–c, osteoderms; (Figure 8 a–l).
Horizon and locality. Cerro del Pueblo Formation (Upper Cretaceous, Campanian), Rincón Colorado locality, General Cepeda, Coahuila, Mexico.
Description. The osteoderms (CPC 3092 a-c) had a dorsal surface with a rise blunt keel orientated caudally, and with and almost flat ventral surface. It has a smooth surface with vascular foramens over their surface. The base is oval in CPC 3092c, except in CPC 3092a and CPC 3092b where the base is slightly broken and only one side is the only preserved edge. Measurements in millimetres of CPC 3092a: length = 31.8; maximum height = 17.5; maximum width = 26.8. CPC 3092b: length = 34.4; maximum height = 21.1; maximum width = 26.9. CPC 3092c: length = 22; maximum height = 14.9; maximum width = 28.5.
Discussion. The three elements were found in close association, that is why were considered as belonging to one individual. The almost flat ventral surface indicates nodosaurian affinity (Coombs & Maryańska, 1990), although its small size cannot be determined if belong to an immature individual or was a small ossicle in a large individual.
4. Discussion
Previous studies indicate the presence of Upper Cretaceous marine deposits at Sierra Mojada (Palomino-Sánchez, pers. comm., 2006). However, the presence of terrestrial fossil vertebrates cast doubts on this interpretation or at least suggests the close proximity of coastal or fluvial environments. The vertebrate fossils described above come from a sequence of sandstones, conglomerates and claystones, which overlie marl beds. The strata above the marls appear to represent fluvial environments, which is supported by the presence of dinosaur bones and fossil tree stumps found in situ (Haarmann, 1913; Janensch, 1926). The palaeoenvironment of the Soledad Beds may have been a coastal plain with marine estuaries that allowed colonization by oysters and sharks (Rivera-Sylva and López-Espinosa, 2006).
Ankylosaur occurrences throughout North America are common in deposits from coastal plains, deltas and in marine shales. So is suggested a preference by ankylosaurs, mainly nodosaurs for coastal habitats bordering the sea. The Soledad Beds may provide additional evidence for latitudinal faunal diversity at that time.
The presence in northern Mexico of Acantholipan gonzalezi, provides evidence for an isolated local endemism, as is also reported for other coeval dinosaurs of Laramidia (Lehman, 1987; 1997; 2001; Sankey, 2008; Rivera-Sylva et al., 2011b; 2017; 2018; Lehman et al., 2019). Late Cretaceous dinosaur assemblages of northern Mexico are taxonomically distinct from coeval assemblages of northern Laramidia. Among ankylosaurs, differentiation around the world is frequently considered to be related to the different types of vegetation consumed by these dinosaurs (Ősi et al., 2016). The diversification of vegetation likely resulted from a north-south climate gradient, with periodic dry to arid intervals that occurred earlier and lasted longer in the south than in the north of Laramidia (Loewen et al., 2013).
Endemism has been suggested for Mexican ankylosaurids, although Euplocephalus sp. has been reported from the southern Aguja Formation (West, 2020), which could be a southern Laramidian species. Endemisms are known from other groups of Campanian dinosaurs (e.g., tyrannosaurids, hadrosaurids, and ceratopsians; Sampson et al., 2004; 2010). The scenario of endemic Mexican ankylosaurs also supports an unknown Late Cretaceous geographical barrier that separated northeastern Mexico from the rest of Laramidia running along the southern shoreline of the continent. This barrier apparently prevented terrestrial faunal interchange (Rivera-Sylva et al., 2011b).
The recent focus on the fossil record of thyreophorans from Mexico has added substantial data for the reconstruction of the biogeography, diversity, and evolutionary patterns of this group of herbivorous dinosaurs. They inhabited a variety of habitats and were opportunistic herbivores. The evolution of ankylosaurs is marked by a progressive widening of the posterior part of the digestive tract, as seen by the increasing aperture of the pelvis (Carpenter, 2012). This change may have been the result of changing diet and the need for a longer fermentation and digestion time.
Ankylosaurids and nodosaurids are known to coexisted in coastal environments, but the dominance of nodosaurids in nearshore deposits in Mexico may indicate a preferred environments along river systems where their carcasses could be more easily transported to the nearby ocean. In contrast, ankylosaurids preferred more inland environments.
CPC 3048 and CPC 3052 were collected from the Cañon del Tule Formation (Upper Maastrichitan), making them the youngest thyreophoran fossil remains recorded in Mexico to date.
5. Conclusions
The diversity of late Cretaceous Mexican ankylosaurs represents a significant episode in the evolution of terrestrial macrovertebrate faunas in Laramidia. The low diversity is probably the result of a bias of the fossil record rather than a real reduction in biological diversity. Although diagnostic material is presently limited, the knowledge on Mexican ankylosaurs will likely improve by the discovery of more complete specimens in the future, as well as by biogeographic comparisons between taxa inhabiting northern and southern Laramidia during the Campanian.
The potential of new discoveries of ankylosaurs from Mexico is very high, and it is likely that many more finds will become available as researches concentrate on the already known localities.
Acknowledgments
We would like to acknowledge Lucia Alfaro-Ortíz (LLF) for taking the photographs and Julio Robledo (MUDE) for their useful skill cleaning the material. Jorge Ortiz-Mendieta (Museo de Historia Natural de Amecameca) is thank for producing figure 1. We also thank Ruben Molina-Pérez for providing the allometric total length scaling of CPC 3046. Additionally, we express our sincere gratitude to Omar Lagarda González and Adolfo Rangel Morelos for their invaluable assistance in creating and contributing to the skeletal reconstruction. We would like to specially thank J. Ignacio Vallejo-González for his invaluable help in the field. We are deeply thankful to Julio Carranza, owner of Rancho Los Ammonites, for allowing access to his property for prospecting We are especially grateful to Ken Carpenter for reviewing a previous version of the manuscript and providing valuable feedback. We also extend our gratitude to the two anonymous reviewers for their valuable comments and suggestions, which greatly improved this manuscript.
References
Arbour, V. M., Burns, M. E., & Sissons, R. L. (2009). A redescription of the ankylosaurid dinosaur Dyoplosaurus acutosquameus Parks, 1924 (Ornithischia: Ankylosauria) and a revision of the genus. Journal of Vertebrate Paleontology, 29 (4), 1117–1135. https://doi.org/10.1671/039.029.0405
Arbour, V. M., & Currie, P. J. (2016). Systematics, phylogeny and palaeobiogeography of the ankylosaurid dinosaurs. Journal of Systematic Palaeontology, 14(5), 385–444. DOI: 10.1080/14772019.2015.1059985
Arbour, V. M., Zanno, L. E., & Gates, T. (2016). Ankylosaurian dinosaur palaeoenvironmental associations were influenced by extirpation, sea-level fluctuations, and geodispersal. Palaeogeography, Palaeoclimatology, Paleoecology, 449, 289–299.
Brown, B. (1908). The Ankylosauridae, a new family of armored dinosaurs from the Upper Cretaceous. Bulletin of the American Museum of Natural History, 24, 187–201.
Brown, C. M., Henderson, D. M., Vinther, J., Fletcher, I., Sistiaga, A., Herrera, J., & Summons, R. E. (2017). An Exceptionally Preserved Three-Dimensional Armored Dinosaur Reveals Insights into Coloration and Cretaceous Predator-Prey Dynamics. Current Biology, 27, 1–8. https://doi.org/10.1016/j.cub.2017.06.071
Brum, A. S., Eleutério, L. H. S., Simões, T. R., Whitney, M. R., Souza, G. A., Sayão, J. M., & Kellner, A. W. A. (2023). Ankylosaurian body armor function and evolution with insights from osteohistology and morphometrics of new specimens from the Late Cretaceous of Antartica. Paleobiology, 49(4), 1–22. https://doi.org/10.1017/pab.2023.4
Burns, M. E., & Sullivan, R. M. (2011). A new ankylosaurid from the Upper Cretaceous Kirtland Formation, San Juan Basin, with comments on the diversity of ankylosaurids in New Mexico. Fossil Record 3. New Mexico Museum of Natural History and Science, Bulletin, 53, 169–178.
Carpenter, K. (1990). Ankylosaur systematics: Example using Panoplosaurus and Edmontonia. In K. Carpenter, & P. J. Currie (eds.), Dinosaur Systematics: Approaches and Presepectives (pp. 281–298). Cambridge University Press.
Carpenter, K. (2012). Ankylosaurs. In M. Brett-Surman, T. R. Holtz, & J. Farlow (Eds.), The Complete Dinosaur (2nd Ed., pp. 505–525). Indiana University Press.
Carpenter, K., Dilkes, D., & Weishampel, D. B. (1995). The dinosaurs of the Niobrara Chalk Formation (Upper Cretaceous, Kansas). Journal of Vertebrate Paleontology, 15, 275–297.
Carpenter, K., & Kirkland, J. I. (1998). Review of Lower and Middle Cretaceous Ankylosaurs from North America. In S. G. Lucas, J. I. Kirkland, & J. W. Estep (Eds.), Lower and Middle Cretaceous Terrestrial Ecosystems (pp. 249–270). New Mexico Museum of Natural History and Science Bulletin 14.
Carpenter, K., Bartlett, J., Bird, J., & Barrick, R. (2008). Ankylosaurs from the Price River Quarries, Cedar Mountain Formation (Lower Cretaceous), east-central Utah. Journal of Vertebrate Paleontology, 28(4), 1089–1101.
Clarac, F., & Quilhac, A. (2019), The crocodylian skull and osteoderms: A functional exaptation to ectothermy?. Zoology, 132, 31–40. https://doi.org/10.1016/j.zool.2018.12.001
Coombs, W. P. (1978). The families of the ornithischian dinosaur order Ankylosauria. Palaeontology, 21(1),143–170.
Coombs, W. P. (1986). A juvenile ankylosaur referable to the genus Euoplocephalus (Reptilia, Ornithischia). Journal of Vertebrate Paleontology, 6(2), 162–173.
Coombs, W. P., & Maryanska, T. (1990). Ankylosauria. In D. B. Weishampel, P. Dodson, & H. Osmólska (Eds.), The Dinosauria (pp. 456–483) University of California Press.
Currie, P. J., Badamgarav, D., Koppelhus, E. B., Sissons, R., & Vickaryous, M. K. (2011). Hands, feet, and behavior in Pinacosaurus (Dinosauria: Ankylosauridae). Acta Palaeontologica Polonica, 56(3), 489–504.
Ford, T. (2000). A review of ankylosaur osteoderms from New Mexico and a preliminary review of ankylosaur armour. In S. G. Lucas, & A. B. Heckert (Eds.), Dinosaurs of New Mexico (pp. 157–176). New Mexico Museum of Natural History and Science, Bulletin 17.
Ford, T., & Kirkland, J. I. (2001). Carlsbald ankylosaur (Ornithischia, Ankylosauria): An ankylosaurid and not a nodosaurid. In K. Carpenter (Ed.), The Armored Dinosaurs (pp. 239–260). Indiana University Press.
Haarmann, E. (1913). Geologische Streifzüge in Coahuila. Zeitschrft der Deutschen Geologischen Gesellschaft, 65(1), 18–47.
Janensch, V. W. (1926). Dinosaurier-Reste aus Mexiko. Centralblatt für Mineralogie, Geologie und Paläontologie, 1926B, 192–197.
Kappus, E. J., Lucas, S. G., & Langford, R. (2011). The Cerro de Cristo Rey Cretaceous Dinosaur tracksites, Sunland Park, New Mexico, USA, and Chihuahua, Mexico. In R. M. Sullivan, S. G. Lucas, & Justin A. Spielmann (Eds.) Fossil Record 3 (pp. 272–288). New Mexico Museum of Natural History and Science, Bulletin 53.
Kirkland J., Alcalá, L., Loewen, M. A., Espílez, E., Mampel, L., & Wiersma, J. P. (2013). The basal nodosaurid ankylosaur Europelta carbonensis n. gen., n. sp. from the Lower Cretaceous (Lower Albian) Escucha Formation of Northeastern Spain. PLoS ONE, 8(12), e80405. https://doi.org/10.1371/journal.pone.0080405
Lehman, T. M. (1987). Late Maastrichtian paleoenvironments and dinosaur biogeography in the Western Interior of North America. Palaeogeography, Palaeoclimatology, Palaeoecology, 60, 189–217.
Lehman, T. M. (1997). Late Campanian dinosaur biogeography in the Western Interior of North America. In D. A. Wolberg, & E. Stump, E. (Eds.), Dinofest International Proceedings (pp. 223–240). Academy of Natural Sciences, Philadephia.
Lehman, T. M. (2001). Late Cretaceous Dinosaur Provinciality. In D. H. Tanke, & K. Carpenter (Eds.), Mesozoic Vertebrate Life (pp. 310–328). Indiana University Press.
Lehman, T. M., Wick, S., Brink, A., Shiller, T. (2019). Stratigraphy and vertebrate fauna of the lower shale member of the Aguja Formation (lower Campanian) in West Texas. Cretaceous Research, 99, 291–314. https://doi.org/10.1016/j.cretres.2019.02.0282
Loewen, M. A., Burns, M. E., Getty, M. A., Kirkland, J. I., & Vickaryous, M. K. (2013). Review of Late Cretaceous Ankylosaurian Dinosaurs from the Grand Staircase Region, Southern Utah. In A. L. Titus, & M. A. Loewen (Eds.), At the top of the Grand Staircase: The Late Cretaceous of Southern Utah (pp. 445–462). Indiana University Press.
Marsh, O. C. (1890). Additional characters of the Ceratopsidae, with notice of new Cretaceous dinosaurs. American Journal of Science series, 3, 39, 81–86.
Martínez-Díaz, J. L., Montellano-Ballesteros, M. (2011). Anquilosaurios (Thyreophora-Ankylosauria) del Cretácico Superior de México. Libro del Resúmenes del XII Congreso Nacional de Paleontología, Puebla, pp. 93.
Meyer, C., Frey, E. D., Thüring, B., Etter, W., Stinnesbeck, W. (2005). Dinosaur tracks from the Late Cretaceous Sabinas Basin (Mexico). Kaupia, 14, 41–45.
Morris, W. J. (1967). Baja California: Late Cretaceous dinosaurs. Science, 155(3769), 1539–1541.
Nopcsa, F. (1915). Die Dinosaurier der siebenbürgischen Landesteile Ungarns. Mitteilungen Jarbuch der Königlich ungarischen geologischen Reichsanstalt, 23, 1–26.
Osborn, H. F. (1923). Two Lower Cretaceous dinosaurs of Mongolia. American Museum Novitates, 95, 1–10.
Ősi, A., & Pereda-Suberbiola, X. (2017). Notes on the pelvic armor of European ankylosaurs (Dinosauria: Ornithischia). Cretaceous Research, 75, 11–22. doi: https://doi.org/10.1016/j.cretres.2017.03.007
Ősi, A., Prondvai, E., Mallon, J., & Reka Bodor, E. (2016). Diversity and convergences in the evolution of feeding adaptations in ankylosaurs (Dinosauria: Ornithischia). Historical Biology, 29(4), 539–570.
Owen, R. (1842). Report on British Fossils Reptiles, Part II. Report of the Eleventh Meeting of the British Association for the Advancement of Science (pp. 60–204). John Murray publisher.
Penkalski, P. (2014). A new ankylosaurid from the Late Cretaceous Two Medicine Formation of Montana, USA. Acta Palaeontologica Polonica, 59(3), 617–634.
Rivera-Sylva, H. E., & Espinosa-Chávez, B. (2006). Ankylosaurid (Dinosauria: Thyreophora) osteoderms from the Upper Cretaceous Cerro del Pueblo of Coahuila, Mexico. Carnets de Géologie, 6, 1–5. https://doi.org/10.4267/2042/4741
Rivera-Sylva, H. E., & López-Espinoza, J. (2006). Informe de una nueva localidad del Cretácico Tardío en Coahuila, México. Libro de resúmenes del X Congreso Nacional de Paleontología, Ciudad de México, México (p. 130). Sociedad Mexicana de Paleontología 10.
Rivera-Sylva, H. E., Carpenter, K., & Aranda-Manteca, F. J. (2011a). Late Cretaceous Nodosaurids (Ankylosauria: Ornithischia) from Mexico. Revista Mexicana de Ciencias Geológicas, 28(3), 271–278.
Rivera-Sylva, H. E., Frey, E., Guzmán-Gutiérrez, R., Palomino-Sánchez, F., & Stinnesbeck, W. A. (2011b). Deinosuchus riograndensis (Eusuchia: Alligatoroidea) from Coahuila, Mexico. Revista Mexicana de Ciencias Geológicas, 28(2), 267–274.
Rivera Sylva, H. E., & Carpenter, K. (2014). Ornithischian dinosaurs from Mexico. In H. E. Rivera Sylva, K. Carpenter, & E. Frey (Eds), Dinosaurs and other reptiles from the Mesozoic from Mexico (pp. 156–180). Indiana University Press.
Rivera-Sylva, H. E., Frey, E., Stinnesbeck, W. A., Guzmán-Gutiérrez, R., & González-González, A. H. (2017). Mexican ceratopsids: Considerations on their Diversity and Biogeography. Journal of South American Earth Sciences, 75, 66–73.
Rivera-Sylva, H. E., Frey, E., Stinnesbeck, W., Carbot-Chanona, G., Sánchez-Uribe, I. E., Guzmán-Gutiérrez, J. R. (2018). Paleodiversity of Late Cretaceous Ankylosauria from Mexico and their phylogenetic significance. Swiss Journal of Palaeontology, 137, 83–93. https://doi.org/10.1007/s13358-018-0153-1
Rivera-Sylva, H. E., Guzmán-Gutiérrez, J. R., Porras-Músquiz, H., Zapata Jaime, R., García de la Garza, J. P., Guajardo Guajardo, R. D., & Galicia Chávez, M. (2019). Paleobiodiversidad de dinosaurios de la Formación Olmos (Campaniano-Maastrictiano) en Coahuila, México. Memorías del 3er Congreso de Diversidad Biológica. Durango, México, 171–176.
Rivera-Sylva, H. E., Nava-Rodríguez, R. L., & Sánchez-Uribe, I. E. (2021). Dinosaurs from the Late Cretaceous of the Ojinaga Basin in Northeastern Chihuahua, Mexico. Paleontología Mexicana, 10(2), 105–111.
Rodríguez-de la Rosa, R. A., Velasco-de León, M. P., Arellano-Gil, J., & Lozano-Carmona, D. E. (2018). Middle Jurassic ankylosaur tracks from Mexico. Boletín de la Sociedad Geológica Mexicana, 70(2), 379–395.
Sampson, S. D., Loewen, M. A., Roberts, E. T., Smith, J. A., Zanno, L. E., & Gates, T. (2004). Provincialism in Late Cretaceous terrestrial faunas: New evidence from the Campanian Kaiparowits Formation of Utah. Journal of Vertebrate Paleontology, supplement 24, 108A.
Sampson, S. D., & Loewen, M. A. (2010). Unraveling a radiation: A review of the diversity, stratigraphic distribution, biogeography, and evolution of horned dinosaurs (Ornithischia: Ceratopsidae). In M. J. Ryan, B. Chinnery Allgeier, & D. A. Eberth (Eds.), New perspectives on horned dinosaurs (pp. 405–427). Indiana University Press.
Sankey, J. T. (2008). Vertebrate Paleobiology from Microsites, Talley Mountain, Upper Aguja Formation (Late Cretaceous), Big Bend National Park, Texas, USA. In J. T. Sankey, & S. Baszio (Eds.), Vertebrate Microfossils Assemblages (pp. 61–77). Indiana University Press.
Seeley, H. G. (1888). On the Classification of the Fossil Animals Commonly Named Dinosauria. Proceedings of the Royal Society of London, 43, 165–171.
Seidel, M. R. (1979). The osteoderms of the american alligator and their functional significance. Herpetologica, 35(4), 375–380.
Sereno, P. (1986). Phylogeny of the bird-hipped dinosaurs (Order Ornithischia). National Geographic Research, 2, 234–256.
Shiller, T. A. (2017). Stratigraphy and paleontology of Upper Cretaceous-Paleogene strata in northern Coahuila, Mexico [PhD Thesis]. Faculty of Texas Tech University.
Vickaryous, M. K., Maryańska, T., Weishampel, D. B. (2004). Ankylosauria. In: Weishampel, D.B., Dodson, P., Osmólska, H. (Eds.), The Dinosauria (2nd Ed., pp. 363–392). University of California Press.
West, B. N. (2020). Campanian-Maastrichtian ankylosaurs of West Texas [Master Thesis]. Texas Christian University.