An ornithopod trackway from the Albian-Turonian Arcurquina Formation, Arequipa, Peru, and its paleoecological implications
Un rastro de ornitópodo, del Albiano-Turoniano de la Formación Arcurquina, Arequipa, Perú, y sus implicaciones paleoecológicas
Guillermo-Ochoa, Angel A.1 ; Zevallos-Valdivia, Leonardo M.1; Castro-Eguiluz, Camila1; Garcia-Flores, Vilma1; Martinez, Jean-Noël2; Silupú-Cárdenas, Omar Alfredo2;
; Zevallos-Valdivia, Leonardo M.1; Castro-Eguiluz, Camila1; Garcia-Flores, Vilma1; Martinez, Jean-Noël2; Silupú-Cárdenas, Omar Alfredo2;
Sánchez-Alva, Claudia Isabel2; Epiquien-Llaja, José Luis2; Pintado-Abarca, Thalía del Pilar2; Delgado-Quiroz, Lizbeth Mariana3; Rodríguez-de la Rosa, Rubén A.3,*
1 Laboratorio de Paleontología, Facultad de Geología Geofísica y Minas. Universidad Nacional de San Agustín de Arequipa, esquina Avenida Independencia con calle Paucarpata S/N, Arequipa, Perú.
2 Instituto de Paleontología, Universidad Nacional de Piura, Campus Universitario, Urbanización Miraflores, Castilla, Piura, Perú.
3 Unidad Académica de Ciencias Biológicas, Universidad Autónoma de Zacatecas, Calzada Solidaridad, esq. Paseo de la Bufa S/N, Campus II, C.P. 98060, Zacatecas, Mexico.
* rubenrodriguezr@uaz.edu.mx
Abstract
Tracks and trackways of swimming ornithopod dinosaurs are rather rare in the ichnological record; in this paper, a short trackway of a small ornithopod dinosaur is described from Albian-Turonian sediments of the Arcurquina Formation, at Yura District, southern Peru. According to the age and overall footprint morphology, a basal member of Iguanodontia is proposed as the potential trackmaker. The trackway consists of three consecutive tridactyl and mesaxonic footprints, preserved as concave impressions or negative epichnia. The first footprint preserves impressions of digits II to IV with a very low angle of total divarication (7.5°). The second footprint preserves a half-moon shaped plantar impression, with no traces of digits II and IV; however, digit III is observed as an elongate and narrow impression, bearing an acuminate end and a proximal subcircular impression; a sediment mound, behind the impression of digit III is preserved as well. The third footprint preserves the whole impression of digit III, while digits II and IV preserve their most distal ends as sub-triangular impressions. The second step is shortest; this characteristic, as well as the different morphology of the three footprints, can be explained by a scenario in which a small ornithopod dinosaur was in underwater locomotion; however, the action of the waves influenced the locomotion of the producer of the footprints. This finding represents an important addition to the ichnological record of dinosaurs in Peru.
Keywords: Arcurquina Formation, Ornithopoda, Peru, subaqueous locomotion, Yura.
Resumen
Huellas y rastros de dinosaurios ornitópodos nadando son raros en el registro icnológico; en esta publicación, el rastro corto de un dinosaurio ornitópodo pequeño, es descrito de sedimentos del Albiano-Turoniano, de la Formación Arcurquina, en el Distrito de Yura, al sur de Perú. De acuerdo con la edad y a la morfología general de las huellas, un miembro basal de Iguanodontia es propuesto como el productor potencial de las huellas. El rastro se conforma por tres huellas consecutivas; estas son tridáctilas, mesaxónicas y se conservan como impresiones cóncavas, o epicnia negativa. La primera huella conserva impresiones de los dígitos II al IV con un ángulo de divergencia total muy bajo, de tan sólo 7.5°. La segunda huella conserva una zona plantar en forma de medialuna e impresiones de los dígitos II y IV ausentes; sin embargo, está presente la impresión, alongada y estrecha del dígito III, con una impresión distal acuminada y una proximal de contorno sub-circular; un montículo de sedimento se encuentra conservado detrás de la impresión del dígito III. La tercera huella conserva una impresión sub-oval, alongada, del dígito III; mientras que los dígitos II y IV conservan impresiones de su parte más distal, con un contorno sub-triangular. El segundo paso se vuelve más corto; esta característica, así como la morfología diferente de las tres huellas, pueden ser explicadas por un escenario en el que un dinosaurio ornitópodo pequeño se encontraba en locomoción subacuática, no obstante, la acción de las olas influyó en la locomoción del productor de las huellas. Este hallazgo representa una adición importante al registro icnológico de dinosaurios en Perú.
Palabras clave: Formación Arcurquina, locomoción subacuática, Ornithopoda, Perú, Yura.
1. Introduction
The vertebrate footprint record from the Mesozoic of Peru includes supposed dinosaur footprints from the Late Cretaceous sediments of the “Couches Rouges” Formation, in the Cusco Region (Gregory, 1916; Leonardi, 1994; Noblet et al., 1987, 1995). However, Lockley et al. (1999) considered that the footprint-bearing layers were Paleogene in age; thus, the footprints were produced by notungulate and/or litoptern mammals (Lockley et al., 1999). The other non-dinosaurian footprint record, known to date, from the Mesozoic of Peru, are footprints related to the ichnogenus Brachychirotherium; these footprints came from Upper Triassic sandstones of the Sarayaquillo Formation in central Peru (Salas-Gismondi and Chacaltana, 2010).
In regard to dinosaur footprints, an early report includes theropod and sauropod footprints that were discovered in a point on the road between Huacamochal and Usquil, at La Libertad Region (Trottereau, 1964). Jaillard et al. (1993) reported Upper Cretaceous theropod and ornithopod footprints from the Vilquechico Group, Vilque Chico District, Huancané Province, Puno Region. In 2016, Jaimes et al. reported a short trackway of a purported theropod dinosaur, also from the Vilquechico Formation (Jaimes et al., 2016). Large-sized dinosaur footprints were reported from the Lower Cretaceous Carhuaz Formation at the Conococha-Antamina road (Obata et al., 2006); these fooprints were also commented on by Arce (2002). One of the best known tracksites from Peru, is located close to the town of Querulpa Chico and 70 km in the northwest of the city of Arequipa, southern Peru. This site preserves 11 trackways belonging to large and small-sized theropod dinosaurs in sediments interpreted as Lower Cretaceous beds of the Hualhuani Formation (Moreno et al., 2004, 2012); however, assigned by local geologists to the Late Jurassic Gramadal Formation. This tracksite was discovered in 2002 and a thematic touristic park has been created around the site.
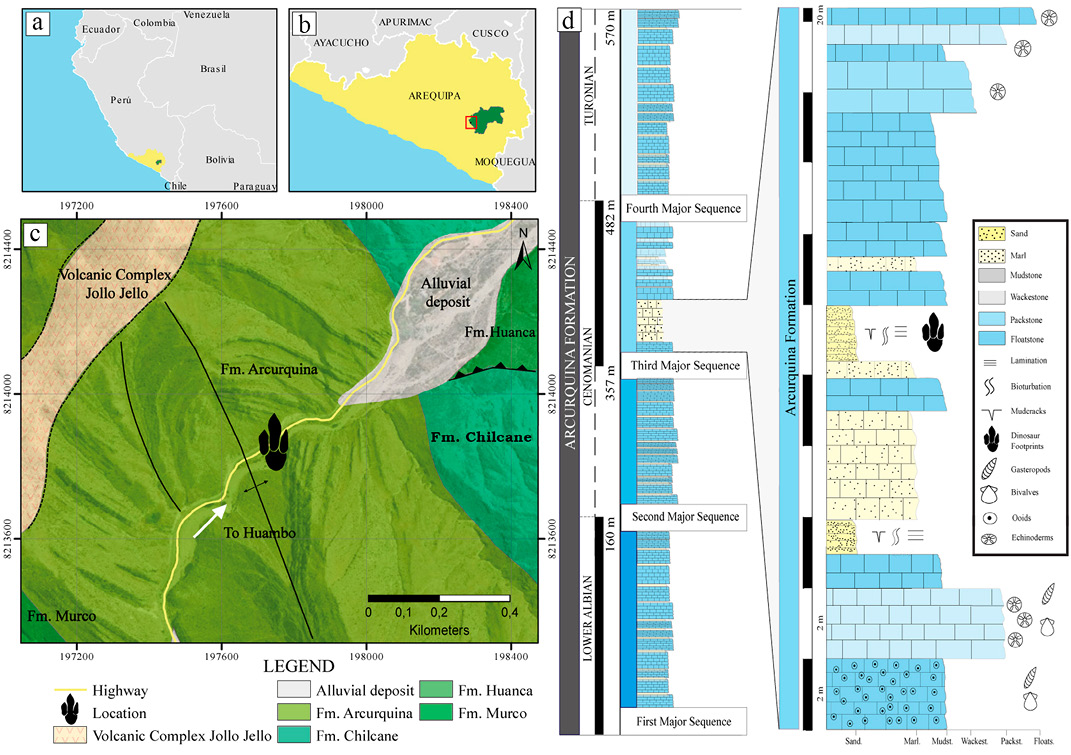
Recently, a short trackway of an ornithopod dinosaur was found in the Albian-Turonian strata of the Arcurquina Formation, at Yura District, in the Province and Region of Arequipa (Figure 1). The footprint morphology suggests a basal member of Iguanodontia in a subaqueous locomotion. It represents the first record of dinosaur footprints in the Arcurquina Formation, Yura District, southern Peru. To report this trackway and its paleoecological implications is the main purpose of this paper.
2. Material and methods
The trackway is preserved in a relatively small outcrop, 2.48 m by 1.40 m; the surface was cleaned manually with brushes. Measurements were taken from the trackway, with a measure tape and a protractor. Divarication angles were measured between the longitudinal axes of the digits. As an additional record of the trackway, footprints were drawn in a plastic sheet with permanent ink markers. Photographs were taken with a Flexacam C5-Leica camera and a Sony Cyber-shot, DSC-WX60 camera.
3. Geological setting
The Arcurquina Formation represents the Albian-Turonian sedimentation in southern Peru, in the regions of Arequipa and Moquegua (Figure 1). In the Yura Basin (Arequipa), it has a thickness of 600 m. Four third order sequences (in terms of sequence stratigraphy) can be recognized in the Arcurquina Formation (Jaillard, 1995). The outcrop consists of a carbonate succession with intermittent detrital and silt-clay episodes (Figure 1d). A stratigraphic correlation with the historical column of Jaillard (1995), suggests that the trackway-bearing outcrop corresponds to sandy beds at the beginning of the third sequence, which are Cenomanian age based on the invertebrate assemblage (Benavides, 1962). The sedimentary environments recorded indicate a shallow to outer carbonate platform, with some lagoon beds and sandy bodies (Jaillard, 1995). The sedimentary facies interpretation corresponding to the third sequence indicates middle to outer ramp facies, interbedded with stratigraphic levels interpreted as tidal flats. The studied trackway is located on a tidal flat level. This layer is composed of coarse to medium-grained sand, with grey and reddish colors due to oxidized allochthonous clasts. It is located at the top of a 3 m small sequence interpreted as tidal flat beds. Mud-cracks sedimentary structures, gypsum cristals and horizontal laminations, are features of this fining upwards sandy sequence. Asymmetric and eroded ripples in the sediment surface (Figure 2a–b) suggests local wave reworking.
In thin section this layer has a lithic composition (quartz 30%, plagioclases 15%, lithic fragments 45%) with a matrix of similar composition (lithic, quartz and chert fragments) and carbonate and siliceous cement (Figure 2c-d). A poorly sorting, low sphericity of grains and reworking ripples can be observed and indicate that these beds represent intertidal environments.
4. Description of the trackway
The reported trackway is conformed by three consecutive footprints preserved in a relatively small outcrop (Figure 3a), two rights and one left; these are preserved as concave impressions, negative epichnia (Figure 3b). Footprints are tridactyl and mesaxonic; however, these show morphological differences, related to the mode of locomotion and the way that the autopodium interacted with the sediment (Figure 3c). The first footprint in the trackway is a right one (Figure 4a); it bears a relatively wide plantar area, with a subcircular heel contour. Its maximum length and width are 23.8 cm and 12.9 cm, respectively. The plantar area is 8.4 cm in length, measured from the posterior area of digit III to the most posterior border of the heel. Digit II has a sub-oval contour, with a rounded proximal end and an acuminate distal end, suggesting the presence of a subtriangular claw. It is 11.0 cm in length and 3.9 cm in maximum width. Digit III bears a conspicuous morphology, with a pyriform-elongate contour, being narrow proximally and wider distally (Figure 4a); it preserves a subcircular proximal border and, distally, a sub-triangular claw impression, 2.0 cm in width, at its base, and 1.4 cm in antero-posterior length. Digit III is 15.4 cm in total length, 4.2 cm in proximal width and 5.6 cm at its widest portion. Digit IV has morphological similarities with digit II; it is sub-oval and elongated, with a rounded proximal end and a distal acuminate end, suggesting the presence of a subtriangular claw. It is 10.2 cm in length and 3.5 cm in maximum width. The angles of divarication between digits, measured from their tips to the heel are 12.5° between digits II and III, and 13.5° between digits III and IV; in this way, the total angle of divarication is 26°. However, the total angle of divarication, measured between the longitudinal axes of digits II and IV, is only 7.5°; it is an important fact that will be discussed later.
The second footprint is a left one (Figure 4b); its morphology varies drastically from the previous right footprint. The plantar area is preserved as a half-moon shaped impression whose apexes represent the contact points with digits II and IV. The total length of this footprint is 21.3 cm and 11.9 cm in maximum width. The plantar area is 9.6 cm, measured from the anterior border (contact with digit III) to the posterior border of the heel. Impressions of digits II and IV are absent. Digit III is elongate, preserved as a “scratch impression”, with a rounded proximal impression, and a distal acuminate end; digit III preserves, as well, a sediment mound of displacement, between its posterior border and the anterior border of the plantar impression (Figure 4b, white arrowhead).
The third footprint, is a right one (Figure 4c), it is tridactyl and mesaxonic; it preserves the whole impression of digit III, and digits II and IV preserve just their distal parts; the wide plantar area observed in previous footprints is absent on this one. This footprint is 16.3 cm in length and 14.2 cm in width, as preserved. Digit III is elongate and suboval in outline, with a roughly acuminate distal end; it is 13.6 cm in length and 7.0 cm at its widest portion. The distal end of the digit II is subtriangular in outline; it is 6.0 cm in length and 6.0 cm at its widest portion. The distal end of the digit IV is pyriform in outline; it is 8.1 cm in length and 4.6 cm at its widest portion. The angle of divarication is 7° between digits II and III, and 10° between digits III and IV; thus, the total angle of divarication (II-IV) is 17°.
The preserved trackway parameters include steps (measured tip to tip of digit III), stride and pace angulation. The step between the first and the second footprint is 34.5 cm. The step between the second and the third footprint is 24.5 cm. The stride measures 63.8 cm (measured between the tips of digit III, of the first and last footprints). The pace angulation is 151°.
5. Discussion
The general morphological features of these footprints suggest that they were produced by the same type of dinosaur; digit III, in particular, is preserved in the first and third footprints, it is suboval and elongated, with a conspicuous robustness (Figure 4); thus, it is suggested that the footprints were produced by the same individual. The presence of a wide plantar area, as well as parallel sided to slightly tapered toes, suggests an ornithopod trackmaker (Thulborn, 1990; Lockley, 1991); according to the age and overall morphology of these footprints, a basal member of Iguanodontia is proposed as the potential trackmaker.
The general morphology of the second footprint (Figure 4b) recalls, somewhat, that of Limayichnus major (Calvo, 1991). In this ichnotaxon, digits II and IV diverge slightly from digit III, at an angle of 25º or less; also, the impression of digit III is oval in shape and separated from the heel impression. As described by Calvo (1991), digits II and IV fuse with the plantar impression, forming a U-shaped structure/impression (Calvo, 1991). A similar configuration is observed in the second footprint from the Yura trackway; however, it lacks the II and IV digit impressions, thus given the half-moon appearance above mentioned. Indeed, Limayichnus major has been related to Iguanodontidae, as well (Calvo, 1991; Calvo and Rivera, 2018). On the basis of the present evidence, no additional ichnotaxonomical assignation is purported for the Yura trackway.
Although each footprint, in the Yura trackway, preserves different morphological traits (Figure 4), the overall morphological features of the footprints are suggestive of an organism in a subaqueous locomotion, as it is explained below. To date, most of the reported non-avian dinosaur swimming traces preserve parallel digit impressions, or “scratch marks”; this is a common feature of dinosaur footprints indicating subaqueous locomotion (e.g., Coombs, 1980; Ezquerra et al., 2007; Navarro-Lorbés et al., 2023). As for example, parallel digit traces are a diagnostic feature in the ichnogenus Characichnos, an ichnogenus related to subaqueous locomotion (Whyte and Romano, 2001; Milner et al., 2006; Xing et al., 2013). According to Navarro-Lorbés et al. (2023), the parallel digit impressions are the result of the, minimal or almost minimal, biomechanical interaction between pes and sediment; this minimal interaction is related to the low influence that it has in the displacement of the animal, being the displacement highly performed by paddling or classical swimming.
As mentioned above, most of the track and trackways of swimming dinosaurs preserve elongate and subparallel digit impressions, ranging from one to, usually, three; also, no trace of the plantar area is preserved. It has been explained by the deepness of the water body crossed by the dinosaur; as the water body got deeper, the dinosaur scratched the bottom with its claws, leaving such subparallel impressions (e.g., Coombs, 1980; Whyte and Romano, 2001; Milner et al., 2006; Ezquerra et al., 2007; Xing et al., 2013; Navarro-Lorbés et al., 2023). An additional feature, observed in footprints indicating subaqueous locomotion, is that most of these parallel “scratch marks”, were left regularly spaced within a particular trackway, as the organism crossed a water body in calm (e.g., Xing et al., 2013). However, Ezquerra et al. (2007) reported an Early Cretaceous swimming dinosaur trackway, from lacustrine nearshore sediments in the Cameros Basin, Spain: the evidence suggests a theropod swimming and scratching the bottom; although the footprints are regularly spaced, amplified asymmetrical walking movements left the right sets of footprints with an inward rotated angle of 40°, in order to maintain direction into a leftward water current (Ezquerra et al., 2007).
In the first footprint of the Yura trackway, the three digits were placed together, almost sub-parallel, with a very low angle of divarication between digits II and IV, just 7.5° (Figure 4a). Thus, it is here assumed that, the almost parallel digit impressions indicate swimming locomotion. In regard to the presence of the plantar impression in the first footprint of the Yura trackway; it is not possible to demonstrate that this footprint is the first one in a trackway of a dinosaur entering in a waterbody; however, it is possible to appreciate the preservation of different footprint morphologies within a single trackway. A similar situation is seen in a Lower Cretaceous tracksite known as “Icnitas-4 tracksite”, at La Rioja, Spain. In this case, the trackway known as EVP12 represents a swimming dinosaur (Perez-Lorente, 2015; Navarro-Lorbés et al., 2023). Although this trackway is composed of five footprints, the morphology of the first one suggests it was made by terrestrial locomotion. However, the second and third footprints (EVP12.2 and EVP12.3; Perez-Lorente, 2015), have elongated and roughly parallel toe impressions, interpreted as the product of the foot slipping in the mud (Perez-Lorente, 2015; Navarro-Lorbés et al., 2023); particularly, the third footprint preserves three elongated digit impressions not convergent in the proximal area (Perez-Lorente, 2015). The explanation of the morphological differences among the footprints from Spain, is that the organism, in this case a dinosaur, entered a water body, from a shallow area to a deeper area (Pieńkowski and Gierliński, 1987; Perez-Lorente, 2015; Navarro-Lorbés et al., 2023).
The second and third footprints of the Yura trackway preserve morphological traits that clearly indicate a swimming locomotion; as mentioned above, the plantar impression in the second footprint is observed as a half-moon structure (Figure 4b); it lacks the impressions of digits II and IV. However, digit III is observed as an elongate and narrow impression, bearing an acuminate distal end and a proximal subcircular impression; a sediment mound, between the impression of digit III and the anterior border of the plantar impression, is preserved as well (Figure 4b, white arrowhead). In this case, it is possible to appreciate that the plantar impression together with digit III participated in the propulsion of the organism.
Structures such as sediment mounds behind digit impressions indicating propulsion have been observed in swimming ornithopod footprints from the Lower Cretaceous Yanguoxia tracksite in China (Fujita et al., 2012: fig. 7e) and from the Lower Cretaceous “Icnitas-4 tracksite”, in Spain; in this later case, the left footprint EVP12.5, from the EVP12 trackway, preserves a large rounded mound of sediment, just behind the toe impressions (Perez-Lorente, 2015: fig. 3.105; Navarro-Lorbés et al., 2023).
The third footprint of the Yura trackway preserves subtriangular impressions, anteroposteriorly short, of the tips of digits II and IV, and a complete impression of digit III (Figure 4c). The overall morphology of this footprint suggests that propulsion was given by digit III and distal portions of digits II and IV. The morphology of this third footprint recalls the footprint morphology of small ornithopods from the famous Cretaceous (Albian-Cenomanian) Lark Quarry tracksite, in the Winton Formation, Queensland, Australia (Romilio et al., 2013; Thulborn, 2016). Some of these ornithopod footprints preserve a prominent digit III, while digits II and IV are observed as sub-triangular impressions; it has been suggested that these footprints were left by swimming ornithopods (Thulborn, 2016: fig. 3).
Among saurischian dinosaurs, swimming traces are well known for theropods (Milner et al., 2006; Whyte and Romano, 2001; Milner and Lockley, 2016). Although questioned by some authors (e.g., Vila et al., 2005; Ishigaki and Matsumoto, 2009; Falkingham et al., 2011, 2012), manus-only and manus-dominated trackways have been associated with swimming sauropods (Bird, 1944; Ishigaki, 1989; Demathieu et al., 2002; Farlow et al., 2019). Tracks and trackways of swimming ornithopod dinosaurs are rather rare in the ichnological record; for example, manus-only trackways attributed to swimming thyreophorans have been reported for ankylosaurids (Riguetti et al., 2021) and suggested for stegosaurians (Romano and Whyte, 2015). In regard to large tracks and trackways of swimming ornithopods, these have been reported by Fujita et al. (2012) and Moklestad et al. (2018); and those of small swimming ornithopods have been reported by Lockley and Foster (2006) and Romilio et al. (2013).
The parameters of the trackway are a notable feature, the step between the first and second footprints is 34.5 cm, the next step between the second and third footprints is shorter, 24.5 cm. The shortening of the second step, as well as the different footprint morphologies in the Yura trackway, could be explained by the scenario of a small ornithopod dinosaur in subaqueous locomotion, however, at the mercy of the waves (Figure 5).
6. Conclusion
This paper reports the first record of dinosaur footprints in the Albian-Turonian Arcurquina Formation and, indeed, from the Yura District, southern Peru. Additionally, it represents a rare finding of a trackway of a small ornithopod, probably a basal member of Iguanodontia, in a subaqueous locomotion being influenced by the waves on the environment. This record is an important addition to the ichnological record of dinosaurs in Peru.
Acknowledgments
Authors acknowledge the support given by the Facultad de Geología, Geofísica y Minas of the Universidad Nacional de San Agustín de Arequipa. RARR and LMDQ thank for their kind support to Mr. Luis Gregorio García Miranda, also to Universidad Autónoma de Zacatecas and Unidad Académica de Ciencias Biológicas, to M.C. Rosa Gabriela Reveles Hernández, Dr. Hans Hiram Pacheco and Dr. Rubén Ibarra Reyes. LMDQ thanks to Professor Mario Adrián Reyes Santana and Diana Laura Sánchez Domínguez (Ayuntamiento 2021-2024 de Genaro Codina, Zacatecas) for their unvaluable support. There is an implicit “thank you” to the entire Mexico-Piura-Arequipa team. This research work was funded by the National Council of Science, Technology and Technological Innovation (CONCYTEC) and the National Program for Scientific Research and Advanced Studies (PROCIENCIA) within the framework of the contest “E041-2024-03 Basic Research Projects” [number of contract PE501087366-2024].
References
Arce, S. (2002). Sobre unas ignitas de dinosaurios en la sierra norte del Perú. Boletín de Lima, 130, 7–10.
Benavides, V. (1962). Estratigrafía pre-Terciaria de la región de Arequipa. Boletín de la Sociedad Geológica del Perú, 38, 5–63.
Bird, R. T. (1944). Did Brontosaurus ever walk on land?. Natural History, 53, 60–67.
Calvo, J. O. (1991). Huellas fósiles de dinosaurios en la Formación Río Limay (Albiano-Cenomaniano), Picún Leufú, Provincia del Neuquén, Argentina (Ornithischia-Saurischia: Sauropoda-Theropoda). Ameghiniana, 28(3–4), 241–258.
Calvo, J. O., & Rivera, C. (2018). Huellas de dinosaurios en la costa oeste del embalse Ezequiel Ramos Mexía y alrededores (Cretácico Superior, Provincia de Neuquén, República Argentina). Boletín de la Sociedad Geológica Mexicana, 70(2), 449–497. http://dx.doi.org/10.18268/BSGM2018v70n2a11
Coombs, W. P. (1980). Swimming ability of carnivorous dinosaurs. Science, New Series, 207(4436), 1198–1200.
Coria, R. A., & Salgado, L. (1996). A basal iguanodontian (Ornithischia: Ornithopoda) from the Late Cretaceous of South America. Journal of Vertebrate Paleontology, 16(3), 445–457.
Demathieu, G., Gand, G., Sciau, J., Freytet, P., & Garric J. (2002). Les traces de pas de dinosaures et autres archosaures du Lias inférieur des Grands Causses, sud de la France. Palaeovertebrata, 31, 1–143. https://doi.org/10.18563/pv.31.1-4.1–143
Ezquerra, R., Doublet, S., Costeur, L., Galton, P. M., & Pérez-Lorente, F. (2007). Were non-avian theropod dinosaurs able to swim? Supportive evidence from an Early Cretaceous trackway, Cameros Basin (La Rioja, Spain). Geology, 35(6), 507–510. https://doi:10.1130/G23452A.
Falkingham, P. L., Bates, K. T., Margetts, L., & Manning, P. L. (2011). Simulating sauropod manus-only trackway formation using finite-element analysis. Biology Letters, 7, 142–145.
Falkingham, P. L., Bates, K. T., & Mannion, P. D. (2012). Temporal and palaeoenvironmental distribution of manus- and pes-dominated sauropod trackways. Journal of the Geological Society, London, 169(2012), 365–370. https://doi: 10.1144/0016-76492011-019
Farlow, J. O., Galton, P. M., & Hyatt, J. A. (2019). Dinosaur footprints from Dinosaur State Park (East Berlin Formation, Lower Jurassic, Rocky Hill, Connecticut, USA). Hallesches Jahrbuch für Geowissenschaften/Beiheft, 46, 27–29.
Fujita, M., Lee, Y.-N., Azuma, Y., & Li, D. (2012). Unusual tridactyl trackways with tail traces from the Lower Cretaceous Hekou Group, Gansu Province, China. Palaios, 27, 560–570. https://doi.10.2110/palo.2012.p12-015r
Gregory, H. (1916). Geological reconnaissance of the Cuzco Valley, Peru. American Journal of Science, 4th Series, 2, p. 241.
Ishigaki, S. (1989). Footprints of swimming sauropods from Morocco. In D. D. Gillette & M. G. Lockley (eds.), Dinosaur tracks and traces (pp. 83–86). Cambridge: Cambridge University Press.
Ishigaki, S., & Matsumoto, Y. (2009). Re-examination of manus-only and manus-dominated sauropod trackways from Morocco. Geological Quarterly, 53(4), 441–448.
Jaillard, E. (1995). La sedimentación albiana-turoniana en el sur del Perú (Arequipa-Puno-Putina). Sociedad Geológica del Perú, Volumen Jubilar Alberto Benavides, 135–157.
Jaillard, E., Cappetta, H., Ellenberger, P., Feist, M., Grambast-Fessard, N., Lefranc, J. P., & Sigé, B. (1993). Sedimentology, paleontology, biostratigraphy, and correlation of the Late Cretaceous Vilquechico Group of Southern Peru. Cretaceous Research, 14, 623–661.
Jaimes, S. F., Salas-Gismondi, R., Chacaltana, C., & Augusto, C. (2016). Nuevas icnitas de un dinosaurio terópodo del Cretáceo superior en sedimentitas de la Formación Vilquechico, Perú. XVIII Congreso Peruano de Geología, 18, 16–19.
Leonardi, G. (1994). Annotated Atlas of South America tetrapod footprints (Devonian to Holocene). Publication of the Companhia de Pesquisa de Recursos Minerais.
Lockley, M. G. (1991). Tracking Dinosaurs, a new look at an ancient world. Cambridge University Press.
Lockley, M. G., Ritts, B. D., & Leonardi, G. (1999). Mammal track assemblages from the Early Tertiary of China, Peru, Europe and North America. Palaios, 14, 398–404.
Lockley, M. G., & Foster, J. R. (2006). Dinosaur and turtle tracks from the Morrison Formation (Upper Jurassic) of Colorado National Monument, with observations on the taxonomy of vertebrate swim tracks. In J. R. Foster & S. G. Lucas (Eds.), Paleontology and Geology of the Upper Jurassic Morrison Formation (pp. 193–198). New Mexico Museum of Natural History and Science Bulletin 36.
Milner, A. R. C., Lockley, M. G., & Kirkland, J. I. (2006). A large collection of well-preserved theropod dinosaur swim tracks from the Lower Jurassic Moenave Formation, St. George, Utah. In J. D. Harris, S. G. Lucas, J. A. Spielmann, M. G. Lockley, A. R. C. Milner & J. I. Kirkland (Eds.), The Triassic–Jurassic Terrestrial Transition (pp. 315–328). New Mexico Museum of Natural History and Science Bulletin 37.
Milner, A. R. C., & Lockley, M. G. (2016). Dinosaur swim track assemblages: characteristics, contexts, and ichnofacies implications. In P. L. Falkingham, D. Marty & A. Richter (Eds.). Dinosaur tracks: The next steps (428 pp.). Indiana University Press.
Moklestad, T., Caneer, T., & Lucas, S. G. (2018). The “lost tracks” at Dinosaur Ridge, Colorado, from the base of the Cretaceous (Late Albian-Early Cenomanian) Mowry Shale Member of the Benton Formation, show a swimming(?) ornithopod affected by a current. In S. G. Lucas & R. M. Sullivan (Eds.). Fossil Record (Number 6, pp. 503–511). New Mexico Museum of Natural History and Science Bulletin 79.
Moreno, K., Jacay, J., Chillitupa, L., De la Vera, P., Hone D., & Benton, M. J. (2004). New dinosaur tracksites from Peru: Evidence for a wide distribution of large theropods during the Late Jurassic-Early Cretaceous in South America. ICHNIA, First International Congress on Ichnology, Argentina, Museo Paleontológico Egidio Feruglio.
Moreno, K., De Valais, S., Blanco, N., Tomlinson, A. J., Jacay, J., & Calvo J. O. (2012). Large theropod dinosaur footprint associations in western Gondwana: Behavioural and palaeogeographic implications. Acta Palaeontologica Polonica, 57(1), 73–83.
Navarro-Lorbés, P., Díaz-Martínez, I., Valle-Melón, J. M., Rodríguez-Miranda, A., Moratalla, J. J., Ferrer-Ventura, M., San Juan-Palacios, R., & Torices, A. (2023). Dinosaur swim tracks from the Lower Cretaceous of La Rioja, Spain: An ichnological approach to non-common behaviours. Cretaceous Research, 147(2023), 105516. https://doi.org/10.1016/j.cretres.2023.105516
Noblet, C., Marocco, R., & Delfaud, J. (1987). Analyse sédimentologique des “Couches Rouges” du Bassin Intramontagneux de Sicuani (Sud du Pérou). Bulletin de l’Institut Français d’Études Andines, 16(1–2), 55–78.
Noblet, C., Leonardi, G., Taquet, P., Marocco, R., & Cordova, E. (1995). Nouvelle découverte d'empreintes laissées par des Dinosaures dans la Formation des Couches Rouges (bassin de Cuzco-Sicuani, Sud du Pérou): conséquences stratigraphiques et tectoniques. Comptes Rendus de l'Académie des Sciences, Paris, 320(2), 785–791.
Obata, I., Gomi, A., Collantes, J., Salas-Gismondi, R., Fujii, N., Kato, S., & Matsukawa, M. (2006). Early Cretaceous dinosaur tracksites newly found in Peru. Annual Report of the Fukuda Geological Institute, 7, 29–43.
Perez-Lorente, F. (2015). Dinosaur Footprints and Trackways of La Rioja. Indiana University Press.
Pieńkowski, G., & Gierliński, G. (1987). New finds of dinosaur footprints in Liassic of the Holy Cross Mountains and its palaeoenvironmental background. Prezegląd Geologiczny, 4, 199–205.
Riguetti, F. J., Citton, P., Apesteguía, S., Zacarias, G.G., & Pereda-Suberbiola, X. (2021). New ankylosaurian trackways (cf. Tetrapodosaurus) from an uppermost Cretaceous level of the El Molino Formation of Bolivia. Cretaceous Research, 124(8), 1–16.
Romano, M., & Whyte, M. A. (2015). Could stegosaurs swim? Suggestive evidence from the Middle Jurassic tracksite of the Cleveland Basin, Yorkshire, UK. Proceedings of the Yorkshire Geological Society, 60, 227–233. https://doi.org/10.1144/pygs2015-354
Romilio, A., Tucker, R. T., & Salisbury, S. W. (2013). Reevaluation of the Lark Quarry dinosaur tracksite (late Albian–Cenomanian Winton Formation, central-western Queensland, Australia): no longer a stampede?. Journal of Vertebrate Paleontology, 33(1), 102–120.
Salas-Gismondi, R., & Chacaltana, C. (2010). Huellas de un arcosaurio Triásico en la Formación Sarayaquillo (Cuenca Ene, Perú) y el icnoregistro de vertebrados en el Mesozoico del Perú. Boletín de la Sociedad Geológica del Perú, 104, 41–58.
Thulborn, R. A. (1990). Dinosaur tracks. Chapman Hall.
Thulborn, T. (2016). Behaviour of dinosaurian track-makers in the Winton Formation (Cretaceous, Albian–Cenomanian) at Lark Quarry, western Queensland, Australia: running or swimming?. Ichnos: An International Journal for Plant and Animal Traces, 24(1), 1–18. https://doi 10.1080/10420940.2015.1129326
Trottereau, G. (1964). Contribution à I'étude des Andes Nord-péruviennes. [Unpublished PhD thesis] Stratigraphie, Université de Grenoble.
Vila, B., Oms, O., & Galobart, À. (2005). Manus‐only titanosaurid trackway from Fumanya (Maastrichtian, Pyrenees): further evidence for an underprint origin. Lethaia, 38, 211–218. https://doi.org/10.1080/00241160510013312.
Whyte, M. A., & Romano, M. (2001) A dinosaur ichnocoenosis from the Middle Jurassic of Yorkshire, UK. Ichnos: An International Journal for Plant and Animal Traces, 8(3–4), 223–234. https://doi.10.1080/10420940109380189
Xing, L. D., Lockley, M. G., Zhang, J. P., Milner, A. R. C., Klein, H., Li, D. Q., Persons IV, W. S., & Ebi, J. F., (2013). A new Early Cretaceous dinosaur track assemblage and the first definite non-avian theropod swim trackway from China. Chinese Science Bulletin, 58, 2370–2378. https://doi.10.1007/s11434-013-5802-6.